CHAPTER 60 Abdomen and pelvis: overview and surface anatomy
GENERAL STRUCTURE AND FUNCTION OF THE ABDOMINOPELVIC CAVITY
Although often considered separately, the abdomen and pelvis form the largest effectively continuous visceral cavity of the body. They act together to provide multiple vital functions including: support and protection of the digestive and urinary tracts and internal reproductive organs and their associated neurovascular supplies; transmission of the neurovascular supply to and from the thorax and the lower limb; provision of support and attachment to the external genitalia and access to and from the internal reproductive and urinary organs; provision of accessory muscles of physiological actions such as respiration, defecation, and micturition; support for the spinal column in weight bearing and movement.
MUSCULOSKELETAL FRAMEWORK OF THE ABDOMEN AND PELVIS
The walls of the abdominopelvic cavity consist of five lumbar vertebrae and their intervening intervertebral discs (lying in the posterior midline); three layers of skeletal muscles (transversus abdominis, internal oblique and external oblique) with associated fasciae and skin (lying lateral and anterolateral); a single muscular layer (rectus abdominis) with its associated fascial coverings (lying anterior); the bony ‘bowl’ formed by the walls of the true and false pelvis (ilium, ischium and pubis on each side); the muscles of the pelvic floor and perineum (lying inferiorly); the diaphragm (lying superiorly) (Fig. 60.1).
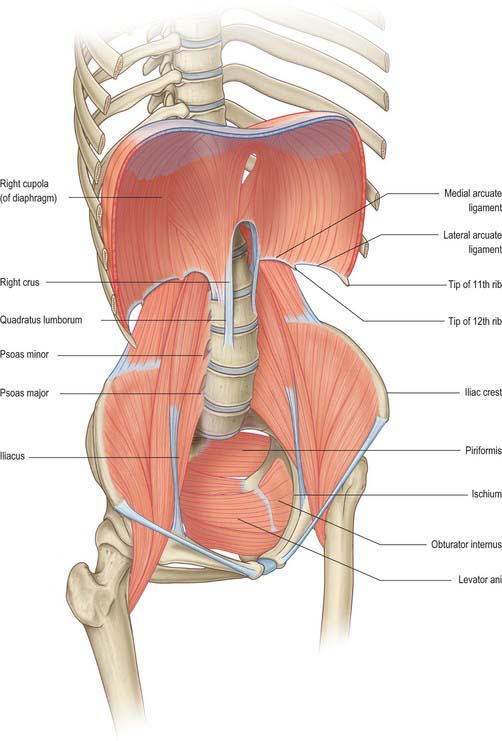
Fig. 60.1 The bony and muscular structures making up the abdominopelvic ‘cavity’. The anterolateral abdominal muscles have been removed for clarity.
The bony protection of the cavity is restricted to the pelvis (true and false) and the upper abdomen, which is partly enclosed by the anterolateral portions of the lower six ribs and their cartilages even though these structures are technically part of the thoracic wall. Between these two zones, the anterolateral abdominal wall is entirely musculofascial but of sufficient thickness and strength that it provides adequate protection for the viscera such that even direct blows can be resisted.
The abdominal wall and retroperitoneal structures play an important role in the function of the spinal column in both movement of the thorax in relation to the pelvis and in aiding support of the spine in weight bearing. The anterolateral muscles provide assistance with flexion and rotation of the thorax in relation to the pelvis (or vice versa if the thorax is fixed).
In the upright position, the pelvis lies at an angle such that the pelvic canal slopes posteriorly at an angle of 45° to the main abdominal visceral cavity. The main abdominal cavity is shaped by the spinal column and the posterior abdominal wall muscles. Although less pronounced than in the thorax, due to the presence of quadratus lumborum and psoas major, its prominence within the abdominal cavity produces two distinct paravertebral gutters on either side of the vertebral column. The lordosis of the lumbar spine in combination with the angle between the fifth lumbar vertebra and the sacrum produces the parabola shape of the sagittal section of each paravertebral gutter.
Thoracoabdominal interface
The thoracic and abdominal cavities interface with each other across the diaphragm. Six principle communications exist between the two cavities: the inferior vena cava, as it crosses between its short supra-hepatic portion and the intrathoracic portion below the right atrium, through the caval opening of the diaphragm (Ch. 62); the oesophagus passing inferiorly through the oesophageal opening of the diaphragm (Ch. 65); the aorta between the descending thoracic and abdominal portions, posterior to the median arcuate ligament of the diaphragm (Ch. 62); the lymphatics of the abdomen draining upwards to the thorax, mainly posterior to the median arcuate ligament via the thoracic duct lying posterolateral to the aorta, but also via peri-caval lymphatics and small vessels draining directly through and at the peripheral insertions of the diaphragm; the azygos and hemiazygos veins ascending behind the median arcuate ligament of the diaphragm into the thoracic azygos system (Ch. 62); the autonomic nervous system, both sympathetic and parasympathetic (see below) which cross between the two cavities behind the medial arcuate ligaments of the diaphragm, via the various diaphragmatic openings, and directly through the substance of the diaphragm itself (Ch. 54). Cutaneous neurovascular structures also cross between the thorax into the abdominal wall structures.
Pelvis–lower limb interface
The pelvis forms an integral part of the bony structure of both the abdominopelvic cavity and the lower limb. It transmits the weight of the upright body from the femoral heads to the lumbar spine as well as providing a stable platform about which movement of the hip joint can occur. Its bony surfaces give rise to extensive muscular attachments for the muscles of the buttock and thigh (Ch. 67) as well as for the muscles of the pelvic floor and perineal diaphragm (Ch. 63). The pelvis also transmits or gives origin to the neuro-lympho-vascular structures which supply the lower limb. The principle structures are as follows. The venous drainage includes the common femoral vein passing under the inguinal ligament to become the external iliac vein; the obturator veins passing through the obturator foramen; the superior and inferior gluteal veins draining from the buttock into the internal iliac vein via the greater and lesser sciatic foramina; multiple smaller venous channels. The arterial supply includes the external iliac artery passing under the inguinal ligament to become the common femoral artery; the obturator artery passing into the thigh via the obturator foramen; the superior and inferior gluteal arteries from the posterior division of the internal iliac artery passing out of the pelvis via the greater and lesser sciatic foramina. The lymphatic drainage principally passes under the inguinal ligament, via the femoral and obturator canals and via the sciatic foramina, to drain into the pelvic lymphatic chains (Ch. 63). The somatic neural structures include the femoral, obturator and sciatic nerves and other minor branches of the lumbosacral plexus (Ch. 62). The autonomic neural structures include those running with the arterial supply and with the branches of the lumbosacral plexus. Cutaneous neurovascular structures also cross between the lower abdomen and pelvis into the skin and superficial tissues of the lower limb.
GENERAL ARRANGEMENT OF ABDOMINOPELVIC NEUROVASCULAR STRUCTURES
It is useful to review here the overall arrangement of the neurovascular systems which cover several regions or multiple viscera of the abdominopelvic region. Descriptions of the neurovascular supply to individual organs are given in the relevant chapters.
AUTONOMIC SUPPLY
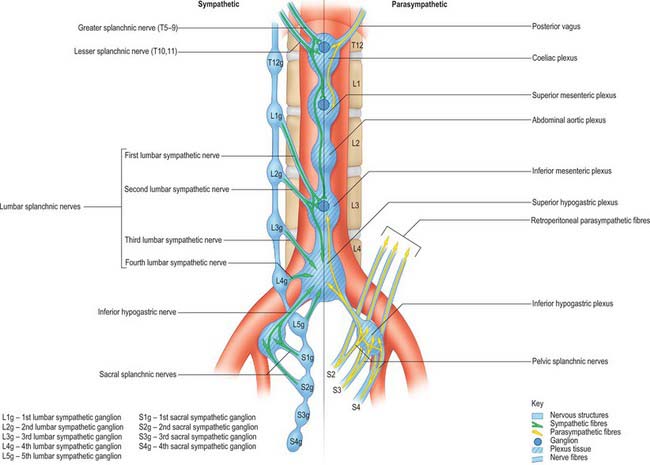
The autonomic supply to the abdominal and pelvic viscera is via the abdominopelvic part of the sympathetic chain and the greater, lesser and least splanchnic nerves (sympathetic), and the vagus and pelvic parasympathetic nerves (parasympathetic). Considerable ramification occurs between plexuses and ganglia, particularly in the major plexuses around the abdominal aorta, hence the descriptions tend to be simplifications based on the ‘main’ supply to each organ (Fig. 60.2). The details of the terminations of these fibres are given on page 1050.
Sympathetic innervation
The cell bodies of neurones of the sympathetic supply of the abdomen and pelvis lie in the intermediolateral grey matter of the first to 12th thoracic and first two lumbar spinal segments. These neurones give rise to myelinated axons which travel in the ventral ramus of the spinal nerve of the same level, leaving it via the white ramus communicans to enter a thoracic or lumbar paravertebral ganglion. Visceral branches may exit at the same level or ascend or descend several levels before exiting but leave the ganglia without synapsing in medial (visceral) branches. These give rise to the greater, lesser and least splanchnic nerves, and the lumbar and sacral splanchnic nerves. Axons destined for supply to somatic structures always synapse in the ganglion of the same level and post-ganglionic, unmyelinated axons leave the ganglion via the grey ramus communicans to enter the spinal nerve of the same level.
Greater splanchnic nerve
The greater splanchnic nerve is derived from the medial, visceral branches of the fifth to ninth thoracic ganglia. It enters the abdomen running through the fibres of the ipsilateral crus of the diaphragm and lies on the crus as it runs anteroinferiorly. The main trunk of the nerve enters the superior aspect of the coeliac ganglion.
Lesser splanchnic nerve
The lesser splanchnic nerve is derived from the medial, visceral branches of the 10th and 11th thoracic ganglia. It enters the abdomen running through the lowermost fibres of the ipsilateral crus of the diaphragm or under the medial arcuate ligament and lies on the crus as it runs anteroinferiorly. The trunk of the nerve enters the aorticorenal ganglion and may give branches to the lateral aspect of the coeliac ganglion. It occasionally joins the greater and least splanchnic nerves as a single splanchnic nerve.
Least splanchnic nerve
The least splanchnic nerve is derived from the medial, visceral branches of the 12th thoracic ganglia. It enters the abdomen running with the sympathetic chain under the medial arcuate ligament and runs inferiorly to enter the renal plexus. The trunk of the nerve enters the aorticorenal ganglion and may give branches to the lateral aspect of the coeliac ganglion. It is sometimes part of the lesser splanchnic nerve, when it forms a twig which enters the renal plexus just below the aorticorenal ganglion.
Lumbar sympathetic system
The lumbar part of each sympathetic trunk usually contains four interconnected ganglia. The trunk runs in the extraperitoneal connective tissue anterior to the vertebral column and along the medial margin of psoas major. Superiorly, it is continuous with the thoracic trunk posterior to the medial arcuate ligament. Inferiorly, it passes posterior to the common iliac artery and is continuous with the pelvic sympathetic trunk. On the right side, it lies posterior to inferior vena cava, and on the left it is posterior to the lateral aortic lymph nodes. It is anterior to most of the lumbar vessels, but may pass behind some lumbar veins.
The first, second and sometimes third lumbar ventral spinal rami send white rami communicantes to the corresponding ganglia. Grey rami communicantes pass from all four lumbar ganglia to the lumbar spinal nerves. They are long, and accompany the lumbar arteries round the sides of the vertebral bodies, medial to the fibrous arches to which psoas major is attached. They supply the sympathetic innervation to the lumbar somatomes via these arteries. Four lumbar splanchnic nerves pass as medial branches from the ganglia to join the coeliac, inferior mesenteric (or occasionally abdominal aortic) and superior hypogastric plexuses. The first lumbar splanchnic nerve, from the first ganglion, gives branches to the coeliac, renal and inferior mesenteric plexuses. The second nerve joins the inferior part of the intermesenteric or inferior mesenteric plexus. The third nerve arises from the third or fourth ganglion and passes anterior to the common iliac vessels to join the superior hypogastric plexus. The fourth lumbar splanchnic nerve from the lowest ganglion passes above the common iliac vessels to join the lower part of the superior hypogastric plexus, or the inferior hypogastric ‘nerve’.
Vascular branches
Vascular branches from all the lumbar ganglia join the abdominal aortic plexus. Fibres of the lower lumbar splanchnic nerves pass to the common iliac arteries and form a plexus which extends along the internal and external iliac arteries. Many postganglionic fibres travel in the muscular, cutaneous and saphenous branches of the femoral nerve, supplying vasoconstrictor nerves to the femoral artery and its branches in the thigh. Other postganglionic fibres travel via the obturator nerve to the obturator artery. Considerable uncertainty persists regarding the exact path of the sympathetic nerve supply to the lower limb (Pick 1970). Sympathetic denervation of vessels in the lower limb can be effected by removing or ablating the upper three lumbar ganglia and the intervening parts of the sympathetic trunk; very rarely, this procedure is useful in treating vascular insufficiency of the lower limb.
Pelvic sympathetic system
The pelvic sympathetic trunk lies in the extraperitoneal tissue anterior to the sacrum beneath the presacral fascia. It lies medial or anterior to the anterior sacral foramina and has four or five interconnected sacral ganglia (they often merge to a greater or lesser extent). Above, it is continuous with the lumbar sympathetic trunk and receives preganglionic fibres descending via the lumbar chain from the lower lumbar spinal segments. Below the lowest ganglia, the two trunks converge to unite in the small ganglion impar anterior to the coccyx. Grey rami communicantes pass from the ganglia to the sacral and coccygeal spinal nerves but there are no white rami communicantes. Medial branches connect across the midline and twigs from the first two ganglia, referred to as sacral splanchnic nerves, join the inferior hypogastric plexus or the hypogastric ‘nerve’ to be distributed to the pelvic viscera via a fine network of pelvic nerves (the ‘pelvic plexus’). Small branches form a plexus on the median sacral artery.
Vascular branches
Postganglionic fibres pass through the grey rami communicantes to the roots of the sacral plexus. Those forming the tibial nerve are conveyed to the popliteal artery and its branches in the leg and foot whilst those in the pudendal and superior and inferior gluteal nerves accompany the same named arteries to the gluteal and perineal tissues. Branches may also supply the pelvic lymph nodes.
Preganglionic fibres for the rest of the lower limb are derived from the lower three thoracic and upper two or three lumbar spinal segments. They reach the lower thoracic and upper lumbar ganglia through white rami communicantes and descend through the sympathetic trunk to synapse in the lumbar ganglia.
Parasympathetic innervation
The parasympathetic neurones innervating the abdomen and pelvis lie either in the dorsal motor nucleus of the vagus nerve or in the intermediolateral grey matter of the second, third and fourth sacral spinal segments. The vagus nerves supply parasympathetic innervation to the abdominal viscera with the exception of the colon distal to the mid descending colon, rectum and upper anal canal. The nerves are derived from the oesophageal plexus and enter the abdomen via the oesophageal opening, closely related to the anterior and posterior walls of the abdominal oesophagus, from which they are separated by loose connective tissue. The anterior vagus is mostly derived from the left vagus and the posterior from the right vagus. The nerves supply the intra-abdominal oesophagus and stomach directly. The anterior nerve supplies branches to the hepatic plexus, which innervates the liver parenchyma and vasculature, the biliary tree including the gallbladder, and the structures in the free edge of the lesser omentum. The posterior nerve supplies branches to the coeliac plexus. These fibres frequently constitute the largest portion of the fibres derived from the plexus: they arise directly from the nerve and from the greater posterior gastric nerve and run beneath the peritoneum, deep to the posterior wall of the lesser sac, at the upper limit of the lesser omentum to reach the coeliac plexus. Their synaptic relays with postganglionic neurones are situated in the myenteric (Auerbach’s) and submucosal (Meissner’s) plexuses (see below).
Pelvic splanchnic nerves
Pelvic splanchnic nerves to the pelvic viscera travel in anterior rami of the second, third and fourth sacral spinal nerves. They leave the nerves as they exit the anterior sacral foramina and pass in the presacral tissue as a fine network of branches which are distributed to three locations. Most pass anterolaterally into the network which forms the inferior hypogastric plexus, whence most are distributed as a fine network of nerves (the ‘pelvic plexus’) to the pelvic viscera (alongside the sympathetic pelvic supply). Some join directly with the fibres forming the inferior hypogastric nerves and hence ascend out of the pelvis, some as far as the superior hypogastric plexus. A few run superolaterally in the presacral tissue, over the pelvic brim anterior to the iliac vessels. They pass directly into the tissue of the retroperitoneum and into the posterior aspect of the mesentery of the sigmoid and descending colon.
The pelvic splanchnic nerves are motor to the smooth muscle of the rectum, anus and bladder wall but inhibitory to the vesical sphincter. They supply vasodilator fibres to the erectile tissue of the penis and clitoris and are probably also vasodilator to the testes, ovaries, uterine tubes and uterus. They are visceromotor to the sigmoid and descending colon as far as the splenic flexure and distal transverse colon.
Abdominopelvic autonomic plexuses and ganglia
The abdominopelvic autonomic plexuses are somewhat variable and often fuse or are closely inter-related. The following descriptions recognize their main features (Figs 60.3, 60.4).
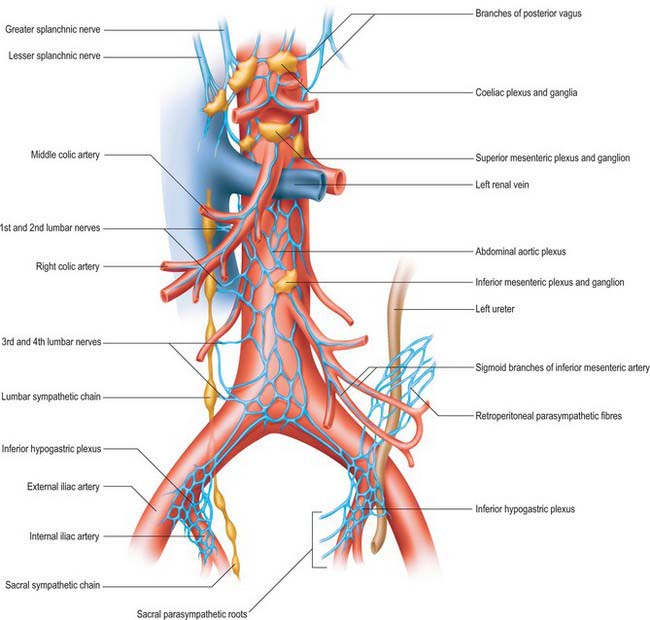
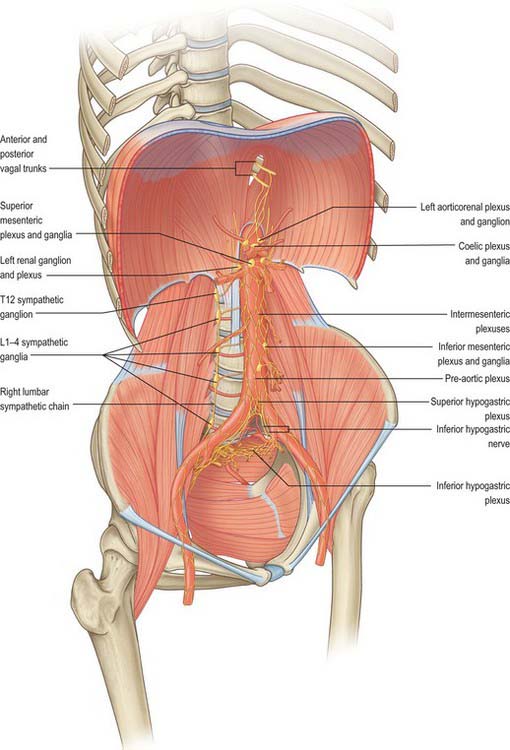
Coeliac plexus
The coeliac plexus is the largest major autonomic plexus, sited at the level of the twelfth thoracic and first lumbar vertebrae. It is a dense network uniting two large coeliac ganglia and surrounds the coeliac artery and the root of the superior mesenteric artery. It is posterior to the stomach and lesser sac, anterior to the crura of the diaphragm and the beginning of the abdominal aorta, and lies between the suprarenal glands. The plexus and ganglia are joined by the greater and lesser splanchnic nerves and branches from the vagus and phrenic nerves. The plexus extends as numerous secondary plexuses along adjacent arteries. The coeliac plexus is connected to, or gives rise to, the phrenic, splenic, hepatic, superior mesenteric, suprarenal, renal and gonadal plexuses (Fig. 60.5).
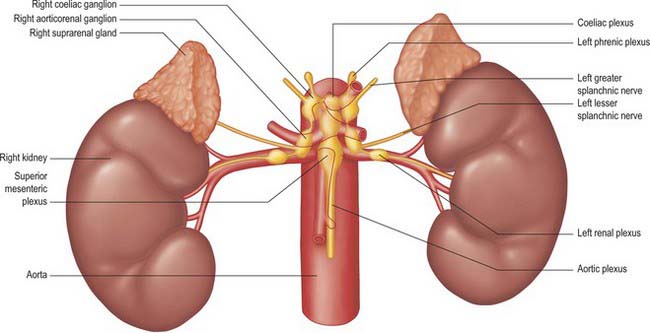
Coeliac and aorticorenal ganglia
The coeliac ganglia are irregular masses on each side of the coeliac trunk adjacent to the suprarenal glands and anterior to the crura of the diaphragm. The right ganglion is posterior to the inferior vena cava, the left ganglion posterior to the origin of the splenic artery. The ipsilateral greater splanchnic nerve joins the upper part of each ganglion. The lower part of each ganglion forms a distinct subdivision, usually termed the aorticorenal ganglion, which receives the ipsilateral lesser splanchnic nerve and gives origin to the majority of the renal plexus (which most commonly lies anterior to the origin of the renal artery).
Phrenic plexus
The phrenic plexus lies around the inferior phrenic arteries on the crura of the diaphragm. It arises as a superior extension of the coeliac ganglion and often receives one or two sensory branches from the phrenic nerve. The left phrenic plexus is usually larger than the right. On the left it supplies branches to the left suprarenal gland and the cardiac orifice of the stomach. The right phrenic plexus joins the phrenic nerve, forming a small phrenic ganglion which distributes branches to the inferior vena cava, suprarenal gland and hepatic plexus.
Superior mesenteric plexus and ganglion
The superior mesenteric plexus lies in the preaortic connective tissue posterior to the pancreas, around the origin of the superior mesenteric artery. It is an inferior continuation of the coeliac plexus, and includes branches from the posterior vagus nerve and the coeliac plexus. Its branches accompany the superior mesenteric artery and divide into secondary plexuses which are distributed along the branches of the artery. The superior mesenteric ganglion lies superiorly in the plexus, usually above the origin of the superior mesenteric artery.
Abdominal aortic plexus (intermesenteric plexus)
The abdominal aortic plexus lies on the sides and front of the aorta, between the origins of the superior and inferior mesenteric arteries. It consists of 4–12 intermesenteric nerves, connected by oblique branches. It is continuous above with the superior mesenteric plexus and below with the superior hypogastric plexus, and is formed by parasympathetic and sympathetic branches from the coeliac plexus. It receives rami from the first and second lumbar splanchnic nerves (which contain sympathetic fibres). The abdominal aortic plexus is connected to the gonadal, inferior mesenteric, iliac and superior hypogastric plexuses.
Inferior mesenteric plexus
The inferior mesenteric plexus lies around the origin of the inferior mesenteric artery and is distributed along its branches. It is formed predominantly from the aortic plexus and the first and second lumbar splanchnic nerves (sympathetic fibres), but it also receives connections from the superior hypogastric plexus (sympathetic and parasympathetic fibres). Disruption of the intermesenteric and inferior mesenteric plexuses rarely causes clinically significant disturbances of autonomic function.
Superior hypogastric plexus
The superior hypogastric plexus lies anterior to the aortic bifurcation, the left common iliac vein, medial sacral vessels, fifth lumbar vertebral body and sacral promontory, and between the common iliac arteries. It is occasionally termed the presacral nerve, but is seldom a single nerve and it is prelumbar rather than presacral. Most frequently found to the left side of the midline, it lies in extraperitoneal connective tissue from which the parietal peritoneum can easily be stripped. The breadth and condensation of its constituent nerves vary. The attachment of the sigmoid mesocolon and upper limit of the mesorectum containing the superior rectal vessels are anterior and to the left of the lower part of the plexus, and separated from it only by a thin sheet of loose connective tissue. The plexus is formed by branches from the aortic plexus and the third and fourth lumbar splanchnic nerves (which are mainly sympathetic). It may also contain parasympathetic fibres from the pelvic splanchnic nerves, which ascend from the two inferior hypogastric plexuses, via a series of filaments which are sometimes identified as the right and left hypogastric ‘nerves’. The latter lie in loose connective tissue just posterolateral to the start of the mesorectum and pass over the pelvic brim medial to the internal iliac vessels. The superior hypogastric plexus supplies fibres to the inferior mesenteric plexus and to the ureteric, gonadal and common iliac plexuses as well as small branches which turn abruptly forward into the upper mesorectum to supply the plexus around the superior rectal artery.
Inferior hypogastric plexus, pelvic plexus and nerves
The inferior hypogastric plexus lies in the thin extraperitoneal connective tissue on the pelvic side-wall anterolateral to the mesorectal fascia. It is directly related to the internal iliac vessels and the attachments of levator ani, coccygeus and obturator internus which lie laterally and the superior vesical and obliterated umbilical arteries superiorly. In males, the inferior hypogastric plexus lies posterolaterally on either side of the seminal vesicles, prostate and the posterior part of the urinary bladder. In females, each plexus lies lateral to the uterine cervix, vaginal fornix and the posterior part of the urinary bladder, and often extends into the broad ligaments of the uterus.
The inferior hypogastric plexus is formed mainly from the sacral splanchnic (sympathetic) and pelvic splanchnic (parasympathetic) branches; a small contribution is made from some sympathetic fibres (from the lower lumbar ganglia) which descend into the plexus via the inferior hypogastric nerves from the superior hypogastric plexus. It gives origin to a complex network of tiny pelvic branches, often referred to as the ‘pelvic plexus’, which supply the pelvic viscera either directly or via periarterial plexuses. The branches of the pelvic plexus supply the vas deferens, seminal vesicles, prostate, accessory glands and penis in males, the ovary, fallopian tubes, uterus, uterine cervix and vagina in females, and the urinary bladder and distal ureter in both sexes. The plexus also gives rise to the long branches of the parasympathetic pelvic splanchnic nerves.
The inferior hypogastric ‘nerves’ are often double, multiple or form an extended plexus on each side. They contain mainly sympathetic and a few parasympathetic fibres. Most of these fibres are ascending from the inferior hypogastric plexus and so it is better to describe them running up out of the pelvis than down into it.
Other plexuses
Separate plexuses exist for organs within the abdomen and pelvis, for example, the hepatic, splenic and gonadal plexuses. Each tends to lie around the main arterial supply to the organ and is supplied with both sympathetic and parasympathetic fibres from one or more of the main plexuses.
Para-aortic bodies
The para-aortic bodies are condensations of chromaffin tissue which are found in close relation to the aortic autonomic plexuses and lumbar sympathetic chains. They are largest in the fetus, become relatively smaller in childhood and have largely disappeared by adulthood. They are most commonly found as a pair of bodies lying anterolateral to the aorta in the region of the intermesenteric, inferior mesenteric and superior hypogastric plexuses. They may lie as high as the coeliac plexus, as low as the inferior hypogastric plexus in the pelvis, or may be closely applied to the sympathetic ganglia of the lumbar chain. Scattered cells which persist into adulthood may, rarely, be the sites of development of a tumour of chromaffin tissue (phaeochromocytoma); these are much more commonly found arising from the cells of the suprarenal medulla. The wide variation in the site of persistent para-aortic body tissue accounts for the range of locations of such tumours.
MAJOR ABDOMINOPELVIC ARTERIAL AND VENOUS ARRANGEMENTS
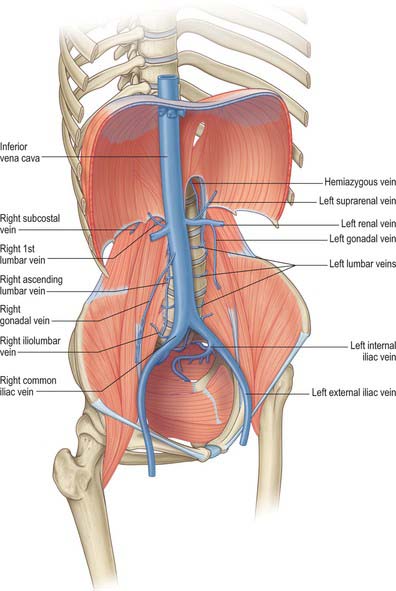
The major vessels which occupy the abdomen and pelvis not only supply the viscera, retroperitoneal structures and much of the bony and soft tissue walls of the cavities, they also course through the cavities en route to supply the lower limbs. Both the arterial and systemic venous trees of the abdomen and pelvis lie predominantly posteriorly in the abdomen and postero-laterally in the pelvis; from the caval and aortic orifices in the diaphragm and the other points where smaller vessels cross the abdominothoracic interface, they follow the general parabolic shape of the lumbar spine. The pelvic divisions follow the contours of the brim and side wall of the true pelvis (Figs 60.6, 60.7). The individual parts of the aorto-iliac and iliocaval systems are described in the appropriate chapters.
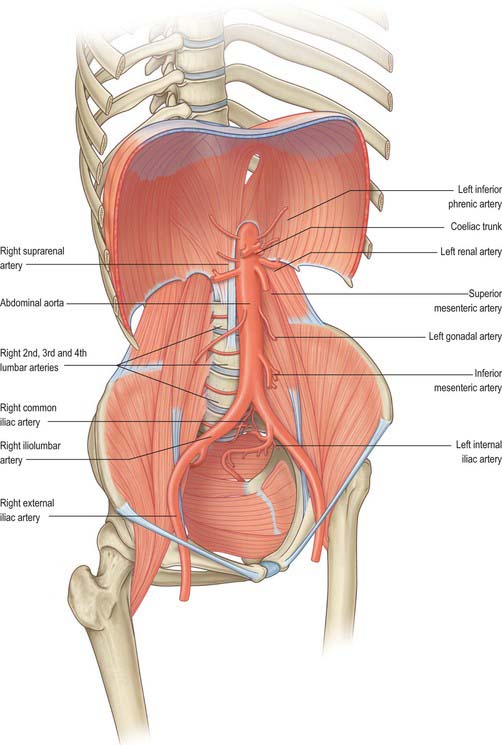
Arterial supply to the gastrointestinal tract
The arterial supply to the gastrointestinal tract is derived from the anterior midline visceral branches of the aorta. There are usually three anterior branches, the coeliac trunk and the superior and inferior mesenteric arteries. Variants in the origin of the arteries are very rare. The most common is a joint origin of the upper two branches as either a coeliacomesenteric trunk or a lienohepaticomesenteric trunk with a separate left gastric artery. Accessory or replaced branches to the upper abdominal viscera are more common (see Chs 65, 66, 67 and 68). Accessory anastomotic vessels between the origin of the coeliac trunk and superior mesenteric artery are rare. The inferior mesenteric artery always arises separately, however, replaced, accessory or anastomotic vessels occasionally arise from the proximal superior mesenteric artery or its branches and contribute to the arterial supply to the proximal territory of the inferior mesenteric artery.
The coeliac trunk and its branches supplies the gastrointestinal tract from the distal third of the oesophagus to the mid part of the duodenum and all derived adenexae (liver, biliary tree, spleen, dorsal pancreas, greater omentum and lesser omentum). The superior mesenteric artery supplies the tract from the mid second part of the duodenum to the distal third of the transverse colon (jejunum, ileum, caecum, appendix, ascending colon and ileal mesentery). The inferior mesenteric artery supplies the tract from the distal transverse colon to the upper anal canal. Other than accessory arteries, numerous medium calibre arteries form anastomoses between the vascular territories. The most profuse occurs around the head of the pancreas and the duodenum, where anastomoses form between the anterior and posterior superior pancreaticoduodenal arteries and the inferior pancreaticoduodenal arteries; the posterior superior pancreaticoduodenal artery and jejunal arteries; the greater pancreatic artery and jejunal arteries. The anastomoses between the territories of the superior and inferior mesenteric arteries are less pronounced and more variable: the most reliable is the pericolic marginal artery, which runs along the transverse colon. Additional anastomotic arcades may exist in the colic mesentery between the transverse and descending colon.
Portal venous system
The (hepatic) portal system, like all portal venous systems, connects two capillary beds: that of the abdominal part of the digestive tube, with the exception of the lower anal canal but including the abdominal part of the oesophagus, and all organs, except the liver, derived from it (i.e. the spleen, pancreas and gallbladder) and the hepatic sinusoidal ‘capillary’ bed. The intrahepatic portal vein ramifies like an artery, and ends in sinusoids from which vessels again converge to reach the inferior vena cava via the hepatic veins. In adults, the portal vein and its tributaries have no valves. In fetal life and for a short postnatal period, valves are demonstrable in its tributaries, but they usually atrophy; although some occasionally persist in an atrophic form into adulthood.
Portal vein
The portal vein begins at the level of the second lumbar vertebra and is formed from the convergence of the superior mesenteric and splenic veins (see Fig. 70.8). It is approximately 8 cm long and lies anterior to the inferior vena cava, posterior to the neck of the pancreas (Fig. 60.8) and obliquely to the right. It ascends behind the first part of the duodenum, the common bile duct and gastroduodenal artery. At this point it is directly anterior to the inferior vena cava. It enters the right border of the lesser omentum, ascends anterior to the epiploic foramen to reach the right end of the porta hepatis, and then divides into right and left main branches which accompany the corresponding branches of the hepatic artery into the liver. In the lesser omentum it lies posterior to both the common bile duct and hepatic artery. The portal vein is surrounded by the hepatic nerve plexus and accompanied by many lymph vessels and some lymph nodes.
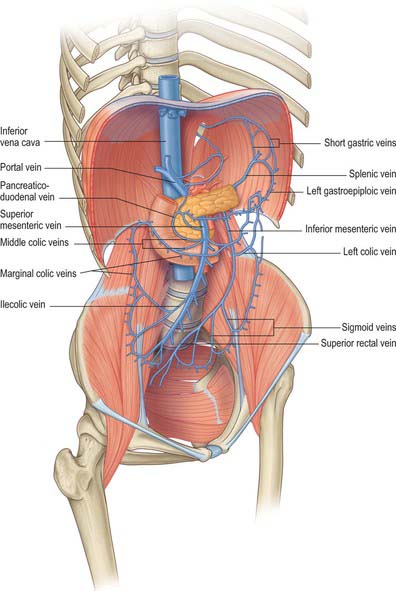
Tributaries of the portal vein
The right branch of the portal vein usually receives the cystic vein and then enters the right lobe of the liver. The left branch receives only the obliterated umbilical vein via the ligamentum teres which connects to its vertical portion. The main extrahepatic tributaries of the main portal vein are the coronary or the left gastric vein which ends in the left margin of the portal vein, and the posterior superior pancreaticoduodenal vein nearer to the head of the pancreas.
GENERAL MICROSTRUCTURE OF THE GUT WALL
The gut wall displays a common structural plan which is modified regionally to take account of local functional differences. The general microstructure is best appreciated by reference to the development of the gut (Ch. 73). Much of the alimentary canal originates as a tube of endoderm enclosed in splanchnopleuric mesoderm. Its external surface faces the embryonic coelom, and the endodermal lining forms the epithelium of the canal and also the secretory and ductal cells of various glands which secrete into the lumen, including the pancreas and liver. The splanchnopleuric mesoderm forms the connective tissue, muscle layers, blood vessels and lymphatics of the wall, and its external surface becomes the visceral mesothelium or serosa. There is no serosa surrounding the cervical and thoracic portions of the gut, or where the hindgut traverses the pelvic floor: in these sites the gut tube is surrounded by a connective tissue adventitia. Neural elements invade the gut from neural crest tissue (Ch. 24). The smooth muscle of the muscularis externa layers of the alimentary canal is supplemented with striated muscle both cranially (from the branchial arches) and caudally. The mature gut wall has four main layers, namely mucosa, submucosa, muscularis externa and serosa (Fig. 60.9). The mucosa (mucous membrane) is the innermost layer and is subdivided into a lining epithelium, an underlying lamina propria (a layer of loose connective tissue, where many of the glands are also found) and a thin layer of smooth muscle, the muscularis mucosae. The submucosa is a strong and highly vascularized layer of connective tissue. The muscularis externa consists of inner circular and outer longitudinal layers which are present throughout the gut wall: a partial oblique layer is present only in the stomach. The external surface is bounded by a serosa or adventitia, depending on its position within the body.
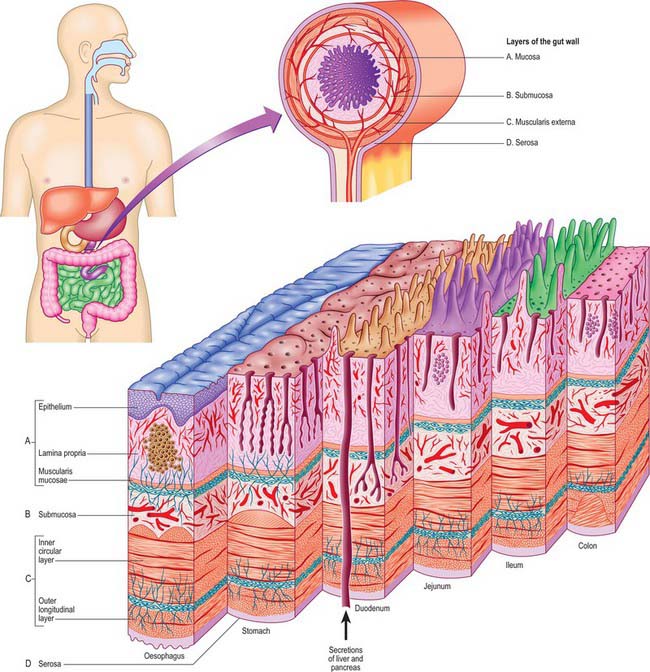
Fig. 60.9 The general arrangement of the alimentary canal to show the layers of the gut wall at the levels indicated.
Mucosa
Epithelium
The epithelium is a protective barrier and the site of secretion and absorption. Its protective function (against mechanical, thermal and chemical injury) is particularly evident in the oesophagus and in the terminal part of the rectum, where it is thick, stratified, and covered in mucus, which serves as a protective lubricant. Other than in these sites, the epithelium lining the gut wall is single-layered, and either cuboidal (in glands) or columnar. It contains cells modified for absorption as well as various types of secretory cell.
The barrier function and selectivity of absorption depend on tight junctions over the entire epithelium. The surface area of the lumen available for secretion or absorption is increased by the presence of mucosal folds, pits, crypts, villi and glands (Fig. 60.9). Microvilli on the surfaces of individual absorptive cells amplify the area of apical plasma membrane in contact with the contents of the gut. Some glands lie in the lamina propria, some in the submucosa, and others (the liver and pancreas) are totally external to the wall of the gut. All of these glands drain into the lumen of the gut through individual ducts. The epithelium also contains scattered neuroendocrine (enteroendocrine) cells.
Lamina propria
The lamina propria consists of compact connective tissue, often rich in elastin fibres, which supports the surface epithelium and provides nutrient vessels and lymphatics. Lymphoid follicles are present in many regions of the gut, most notably in Peyer’s patches. Cells within the lamina propria are the source of growth factors which regulate cell turnover, differentiation and repair in the overlying epithelium.
Muscularis mucosae
The muscularis mucosae is well developed in the oesophagus and in the large intestine, especially in the terminal part of the rectum. In addition, single muscle cells originating from the muscularis mucosae are found inside the villi or between the tubular glands of the stomach and large intestine. By its contraction, the muscularis mucosae can alter the surface configuration of the mucosa locally, allowing it to adapt to the shapes and mechanical forces imposed by the contents of the lumen, and in the intestinal villi, promoting vascular exchange and lymphatic drainage.
Submucosa
The submucosa contains large bundles of collagen and is the strongest layer of the gut wall. However, it is also pliable and deformable and can therefore adjust to changes in the length and diameter of the gut. It contains the largest arterial network of the wall, which supplies both the mucosa and the muscle coat. The submucosa invades the folds which project into the lumen of the oesophagus and rectum, the rugae of the gastric wall, and the plicae circulares of the small intestine, but does not enter the villi.
Muscularis externa
The muscularis externa usually consists of distinct inner circular and outer longitudinal layers whose antagonistic activities create waves of peristalsis responsible for the movement of ingested material through the lumen of the gut. In the stomach, where movements are more complex, there is a partial oblique layer, internal to the other two layers. The layer of circular muscle is invariably thicker than the longitudinal muscle, except in the colon, where the longitudinal muscle is gathered into three cords (taenia coli).
The muscularis externa is composed almost exclusively of smooth muscle, except in the upper oesophagus, where smooth muscle blends with striated muscle. Although the oesophageal musculature resembles that of the pharynx, it is entirely under involuntary control. For most of its length, the smooth muscle of the gut wall consists of ill-defined bundles of cells, typically visceral in type, and somewhat larger than vascular smooth muscle cells. They are approximately 500 μm long, regardless of body size, and are electrically and mechanically coupled. Their fasciculi lack a perimysium, but have sharp boundaries.
The arrangement of the musculature means that a segment of gut can change extensively in diameter (to virtual occlusion of the lumen) and also in length, although elongation is limited by the presence of mesenteries. The co-ordinated activity of the two muscle layers produces a characteristic motor behaviour which is mainly propulsive and directed anally (peristalsis), combined with a non-propulsive motor activity which either mixes the luminal contents, as occurs in the stomach, or partitions them, as occurs at the pyloric sphincter. The muscle maintains constant volume, so that shortening of a segment of the gut wall is accompanied by an increase in muscle layer circumference.
Intestinal smooth muscle exhibits variable and changing degrees of contraction on which rhythmic (or phasic) contractions are superimposed. Slow waves of rhythmic electrical activity, driven by changes in membrane potential in pacemaker cells (interstitial cells), spread throughout the thickness of the circular and longitudinal smooth muscle coats. After spreading circumferentially, slow waves can move in either oral or anal directions, causing segmental contraction. The distances of propagation and the patterns of this spontaneous activity vary between areas of the intestine. Neural regulation of slow and phasic contractions involves excitatory and inhibitory transmitters which are released from the myenteric plexus. This motor control is closely co-ordinated with mucosal absorption and secretion and is mediated via intrinsic nerves in the submucous plexus. The peristaltic reflex occurs during passage of luminal contents down the intestine. It involves ascending contraction and descending relaxation: the sensory limb is mediated by sensory neurones that respond to either mucosal stimulation (intrinsic primary afferents) or muscle stretch (extrinsic afferents).
Interstitial cells
Interstitial cells (of Cajal) are thought to act as pacemakers for the myogenic contraction of muscularis externa, establishing the rhythm of bowel contractions through their influence on electrical slow wave activity. They receive modulatory inputs from the enteric nervous system and from the extrinsic innervation of the gut.
Interstitial cells are thin, flat, and branched. They resemble smooth muscle cells because they contain actin and myosin filaments, and are linked by gap junctions to typical smooth muscle cells. However, they are phenotypically distinct from muscle cells because their intermediate filaments are vimentin rather than desmin, (which is typical of muscle cells). They lie in close apposition to varicose nerve endings of at least two types; one contains small, round clear vesicles (50 nm diameter), the other contains flat, discoidal vesicles (70 nm diameter).
The positions of the interstitial cell layers vary regionally. In general, they lie in the same layers as the enteric plexuses. They are scattered among the cells of the circular muscle layer in the oesophagus and stomach, lie between the inner and outer layers of circular smooth muscle in the small intestine, and colocalize with the myenteric plexus and the single layer of the submucosal plexus on the luminal side of the circular component of the muscularis externa, in the large intestine.
Serosa and adventitia
A layer of connective tissue, of variable thickness, lies external to the muscularis externa. In many places it contains adipose tissue. Where the gut is covered by visceral peritoneum, the external layer is a serosa, which consists of a thin connective tissue layer and an external coat of mesothelium. Elsewhere the connective tissue blends with that of the surrounding fasciae and is referred to as an adventitia. Where the alimentary tract is retroperitoneal, the surface facing the abdominal cavity is covered by serosa, and the other parts are covered by adventitia.
Vascular plexuses
Vascular plexuses are present at various levels of the wall, especially in the submucosa and mucosa: they connect with plexuses of vessels which supply the surrounding tissues or those entering through the mesentery, and accompany the ducts of outlying glands.
Innervation
The gut is densely innervated by the autonomic and enteric nervous systems, and is under extrinsic and intrinsic neuronal control. Neuronal cell bodies of the enteric nervous system lie between the circular and longitudinal components of the muscularis externa (myenteric plexus) and within the submucosa (submucosal plexus). They provide the intrinsic sensory and motor supply of the gut wall and connect with extrinsic sensory, motor and sensorimotor nerves of cranial or spinal origin.
Extrinsic innervation
The extrinsic innervation is derived from neurones outside the gut, and contains functional components from the sympathetic, parasympathetic and visceral sensory divisions of the peripheral nervous system (see p. 1041). Visceral sensory endings respond to excessive muscular contraction or distension: their cell bodies are situated in the nodose ganglion of the vagus nerve and in thoracic and lumbar spinal or dorsal root ganglia. The cell bodies of parasympathetic efferent axons lie in the vagal dorsal motor nucleus in the medulla oblongata. Sympathetic efferent fibres arise from the thoracic and lumbar spinal cord and relay in prevertebral sympathetic ganglia (coeliac, mesenteric and pelvic).
A subserosal plexus, which sometimes contains neuronal cell bodies, connects the extrinsic nerve fibres with the myenteric plexus and is particularly prominent near the mesentery. Fibres from this plexus run through the longitudinal muscle layer to reach the myenteric plexus.
Intrinsic innervation
The intrinsic innervation of the gut wall is derived from neurones which are located entirely within the wall in intramural ganglionated plexuses. The myenteric (Auerbach’s) plexus is a network of fine bundles of axons and small ganglia which lies within the muscularis externa, between the circular and longitudinal layers. It is often associated with secondary and tertiary plexuses of nerve fibres which sometimes contain isolated neuronal cell bodies. There are two or more submucosal plexuses, the most superficial of which is Meissner’s plexus.
Non-ganglionated nerve plexuses lie at various levels in the wall, notably in the lamina propria (mucosal plexus); at the interface between the submucosa and muscularis externa; between the circular and longitudinal muscles (the non-ganglionated part of the myenteric plexus); and within the serosa. An additional non-ganglionated plexus lies between the internal and external components of the circular muscle of the small intestine. All parts of the myenteric plexus are continuous not only with each other, but also with the nerve fibre bundles in the circular muscle. The latter are connected to the ganglionated and non-ganglionated plexuses of the submucosa, and these in turn are connected with the mucosal plexus by fibres which pass through the muscularis mucosa.
SURFACE AND DIAGNOSTIC ANATOMY OF THE ABDOMEN AND PELVIS
ABDOMINAL PLANES AND REGIONS
For descriptive purposes, the abdomen can be divided by a number of imaginary horizontal and vertical lines drawn using the skeletal landmarks of the thorax and abdomen (Fig.). Projection of these lines into the sagittal or transverse planes can then be used to define certain abdominal ‘planes’. Apart from dividing the abdomen into different regions for descriptive purposes, these planes are also of value in defining approximate vertebral levels and the positions of some relatively fixed intra-abdominal structures.
Vertical planes
In addition to the midline, which passes through the xiphisternal process and the pubic symphysis, there are two paramedian planes which are projected from the midclavicular line (also sometimes called the lateral or the mammary line). This line passes through the midpoint of the clavicle, crosses the costal margin just lateral to the tip of the ninth costal cartilage, and passes through a point mid way between the anterior superior iliac spine and the symphysis pubis. It approximates to, but does not exactly correspond to, the lateral border of rectus abdominis.
Horizontal planes
Several horizontal planes have been defined, but only the subcostal and transtubercular planes are in common clinical use.
The xiphisternal plane runs horizontally through the xiphoid processes at the level of the ninth thoracic vertebra. It demarcates the level of the cardiac plateau on the central part of the upper border of the liver.
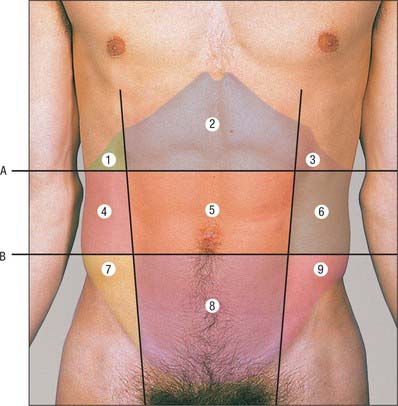
Fig. 60.10 Planes and regions of the abdomen. Key for planes: A, Subcostal plane. B, Transtubercular plane. Key for nine regions of the abdomen: 1. Right hypochondrium. 2. Epigastric. 3. Left hypochondrium. 4. Right lumbar. 5. Central/umbilical. 6. Left lumbar. 7. Right iliac fossa. 8. Suprapubic/hypogastrium. 9. Left iliac fossa.
(Photograph by permission from Lumley JSP 2002 Surface Anatomy, 3rd edn. Edinburgh: Churchill Livingstone.)
The transpyloric plane lies midway between the suprasternal notch of the manubrium and the upper border of the pubic symphysis. It usually lies at the level of the body of the first lumbar vertebra near its lower border and meets the costal margins at the tips of the ninth costal cartilages, where a distinct ‘step’ may be felt at the costal margin. The linea semilunaris crosses the costal margin on the transpyloric plane. The hilum of both kidneys, the origin of the superior mesenteric artery, the termination of the spinal cord, the neck, adjacent body and head of the pancreas, and the confluence of the superior mesenteric and splenic veins as they form the hepatic portal vein may all lie in this plane. The pylorus may be found in the transpyloric plane, but is not a constant feature.
The subcostal plane is a line joining the lowest point of the costal margins, formed by the tenth costal cartilage on each side. It usually lies at the level of the body of the third lumbar vertebra, the origin of the inferior mesenteric artery from the aorta, and the third part of the duodenum, although this varies with posture.
The supracristal plane joins the highest point of the iliac crest on each side. It usually lies at the level of the body of the fourth lumbar vertebra, and marks the level of bifurcation of the abdominal aorta. On the posterior abdominal surface, it is a common level for the identification of the fourth lumbar vertebra, and is used to perform lumbar puncture at the L4–5 or L5–S1 intervertebral level, which is safely below the termination of the spinal cord.
The transtubercular plane joins the tubercles of the iliac crests and usually lies at the level of the body of the fifth lumbar vertebra near its upper border. It indicates, or is just above, the confluence of the common iliac veins and marks the origin of the inferior vena cava.
The interspinous plane joins the centres of the anterior superior spines of the iliac crests. It passes through either the lumbosacral disc, or the sacral promontory, or just below them, depending on the degree of lumbar lordosis, sacral inclination and curvature.
The plane of the pubic crest lies at the level of the inferior end of the sacrum or part of the coccyx, depending on the degree of lumbar lordosis, sacral inclination and curvature.
Abdominal regions
The abdomen can be divided into nine arbitrary regions by the subcostal and transtubercular planes and the two midclavicular planes projected onto the surface of the body. These regions are used in practice for descriptive localization of the position of a mass or the localization of a patient’s pain, and may also be used in the description of the location of the abdominal viscera. The nine regions thus formed are: epigastrium; right and left hypochondrium; central or umbilical; right and left lumbar; hypogastrium or suprapubic; right and left iliac fossa.
ANTERIOR ABDOMINAL WALL
Skeletal landmarks
The superior boundary of the anterior abdominal wall is formed by several clear landmarks (Fig. 60.11). In the midline superiorly lies the xiphoid process. From this point, the costal margins extend to either side from the seventh costal cartilage at the xiphisternal joint to the tip of the twelfth rib (the latter is often difficult to feel in the obese or if it is short). The lowest part of the costal margin lies in the midaxillary line and is formed by the inferior margin of the tenth costal cartilage. The tip of the lower border of the ninth costal cartilage can usually be defined as a distinct ‘step’ along the costal margin.
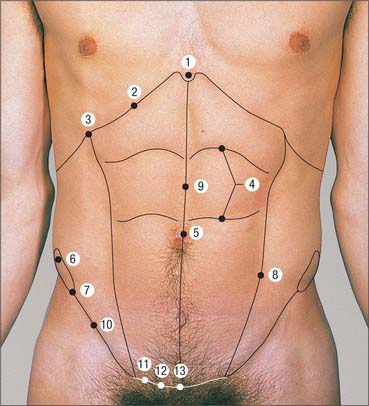
Fig. 60.11 Anterior abdominal wall landmarks. 1. Xiphoid process. 2. Costal margin. 3. Tip of the ninth costal cartilage. 4. Tendinous intersections. 5. Umbilicus. 6. Iliac crest. 7. Anterior superior iliac spine. 8. Linea semilunaris. 9. Linea alba. 10. Inguinal ligament. 11. Pubic tubercule. 12. Pubic crest. 13. Pubic symphysis.
(Photograph by permission from Lumley JSP 2002 Surface Anatomy, 3rd edn. Edinburgh: Churchill Livingstone.)
The inferior boundary of the anterior abdominal wall is formed, in order, by the iliac crest, which descends from the tubercle of the iliac crest to the anterior superior iliac spine; the inguinal ligament, which runs downwards and forwards to the pubic tubercle; and the pubic crest, which runs from the pubic tubercle laterally to the pubic symphysis in the midline. The pubic tubercle can be identified by direct palpation in thin individuals and can be detected, even in the obese, by palpation of the tendon of adductor longus, which runs up to its attachment on the pubis directly below the pubic tubercle. The tendon is best felt in tension with the hip flexed, abducted and externally rotated.
The posterolateral boundary is defined by the midaxillary line.
Soft tissue landmarks
Umbilicus
The umbilicus is an obvious but very inconstant landmark. In the supine adult, it usually lies at the level of the disc between the third and fourth lumbar vertebrae. The bifurcation of the abdominal aorta then lies about 2 cm caudal to the umbilicus. In the erect position, in children and in the obese, or in individuals with a pendulous abdomen, the umbilicus may lie at a lower level.
Rectus abdominis
In a thin and muscular individual, the tendinous intersections of rectus abdominis may be visible when the muscle is tensed by lifting the head against resistance or by sitting up: the intersections are usually situated at the level of the umbilicus, the level of the xiphoid process and mid way between these two points.
Linea alba
The linea alba is usually only visible in thin muscular individuals. It is wide and obvious above the umbilicus, but is almost linear and invisible below this level.
Linea semilunaris
The linea semilunaris lies along the lateral margin of the rectus sheath and is visible as a shallow curved groove in muscular individuals when the abdominal muscles are tensed, e.g. by sitting up from the lying position.
Inguinal region
The two commonly described surface markings in the inguinal region are the mid-inguinal point and the midpoint of the inguinal ligament.
The mid-inguinal point lies at the midpoint of the line between the symphysis pubis and the anterior superior iliac spine. It is superior to the point of palpation of the pulse of the femoral artery as it emerges from beneath the inguinal ligament. The approximate surface marking of the deep inguinal ring lies immediately above the mid-inguinal point, and is a useful landmark for palpation of the origin of an indirect inguinal herniae sac.
INTRA-ABDOMINAL VISCERA
The surface markings of the intra-abdominal viscera are variable and depend on age, body habitus, nutritional state, phase of ventilation and body position. The use of radiological imaging of the abdominal viscera means that the location of viscera by surface markings is almost obsolete in modern clinical practice. The following descriptions are at best regarded as the most common or approximate markings in a healthy supine individual (Fig. 60.12).
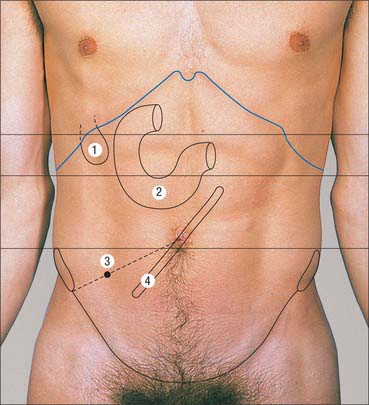
Fig. 60.12 Intra-abdominal visceral landmarks. 1. Gallbladder. 2. Duodenum. 3. Appendix. 4. Root of small bowel mesentery.
(Photograph by permission from Lumley JSP 2002 Surface Anatomy, 3rd edn. Edinburgh: Churchill Livingstone.)
Stomach
The stomach lies in a curve within the left hypochondrium and epigastrium although, when distended and pendulous, it may lie as far down as the central or hypogastric regions. The epigastrium is the usual place to auscultate for a ‘succussion splash’ caused by chronic gastric stasis in upper intestinal obstruction.
Duodenum
The first part of the duodenum sometimes lies just above the trans-pyloric plane, depending on its mobility and length. The second part usually lies in the transpyloric plane just to the right of the midline, and the third part usually lies in the subcostal plane across the midline. The fourth part often lies in the transpyloric plane to the left of the midline, although its position varies according to the length of its mesentery.
Small bowel and its mesentery
The small bowel is usually referred to as lying in the central umbilical region, but often occupies part of both iliac fossae, both lumbar regions and the hypogastrium. The small bowel mesentery runs obliquely in a line from a point 2 cm to the left of the body of the second lumbar vertebra above the subcostal plane, to a point just anterior to the right sacroiliac joint just below the interspinous plane.
Appendix
The appendix is highly variable both in its length and in its position. Its base is commonly referred to as lying beneath a point marked two-thirds of the way along a line joining the umbilicus to the anterior superior iliac spine.
Liver
The inferior border of the liver extends along a line that passes from the right 10th costal cartilage to the left fifth rib in the midclavicular line. The upper border of the liver follows a line that passes from the fifth rib in the midclavicular line on the right to the equivalent point on the left. This upper border curves slightly downwards at its centre and crosses the midline behind the xiphoid. The right border of the liver is curved to the right and joins the upper and lower right limits. The outline of the liver may be defined by the dull note it gives on percussion when compared with the resonance of the lungs above and the hollow abdominal viscera below.
The lower edge of a normal liver cannot usually be palpated, even in women and children, in whom the liver is at a slightly lower level. What is often mistaken for the liver in abdominal palpation is the bulge of rectus abdominis above its upper tendinous intersection.
Gallbladder
The fundus of the gallbladder is very variable in location. It is commonly identified with the tip of the ninth costal cartilage (in the transpyloric plane), near the junction of the linea semilunaris with the costal margin.
Spleen
The spleen lies beneath the ninth, 10th and 11th ribs on the left side. Its surface markings can be delineated on the lower posterior thoracic wall by defining its axis, which extends from a point 5 cm to the left of the midline at the level of the 10th thoracic spine, and passes laterally along the line of the tenth rib to the midaxillary line.
RETROPERITONEAL VISCERA
The surface projections of the retroperitoneal viscera are fairly reliable but have little clinical use since most interventions and procedures are guided by radiological imaging.
Pancreas
The surface projection of the head of the pancreas lies within the duodenal curve. The neck lies in the transpyloric plane, behind the pylorus in the midline. The body passes obliquely up and to the left for approximately 10 cm, its left part lying a little above the transpyloric plane. The tail lies a little above and to the left of the intersection of the transpyloric and left lateral planes.
Kidney
The anterior and posterior surface projections of the kidneys are related to anterior and posterior abdominal wall landmarks. In the description that follows, all measurements are approximate. The right kidney lies 1.25 cm lower than the left. On the anterior surface, the centre of the hilum lies in the transpyloric plane 5 cm from the midline and slightly medial to the tip of the ninth costal cartilage. From the hilum, the outline of the anterior surface can be drawn 11 cm long and 4.5 cm broad, the upper pole being 2.5 cm, and the lower 7.5 cm, from the midline: since the transverse axis is oblique, the width thus shown is 1.5 cm less than the actual width of the kidney. On the posterior surface, the centre of the hilum lies opposite the lower border of the spinous process of the first lumbar vertebra and 5 cm from the midline. The outline of the posterior surface can be traced similarly to the anterior surface. The lower pole is 2.5 cm above the summit of the iliac crest. The kidneys are 2.5 cm lower in the standing than in the supine individual; they ascend and descend a little with respiration, which is of particular importance in endoscopic renal surgery. The lower pole of the normal right kidney may occasionally be felt in the thin individual, especially the female, by bimanual palpation on full inspiration.
Ureter
The ureter starts on either side approximately at the transpyloric plane (the left higher than the right), approximately 5 cm from the midline. Each passes downwards and somewhat medially to enter the bladder at a point marked superficially by the position of the pubic tubercle.
Abdominal aorta
The abdominal aorta starts just to the left of the midline, at the level of the body of the 12th thoracic vertebra. It continues downwards for 10 cm as a band 2 cm wide, and bifurcates at the level of the fourth lumbar vertebra (which is marked by the transtubercular plane), 1.5 cm below and to the left of the umbilicus. The pulsations of the aorta can be felt in a thin individual in the supine position by pressing firmly in the midline backwards onto the vertebral column. An easily palpable aorta in an obese person should raise the suspicion of an aneurysm, to be checked by radiological imaging.
Visceral arteries
The coeliac trunk arises from the aorta immediately after it enters the abdomen at the level of the 12th thoracic vertebra. The origin of the superior mesenteric artery usually lies above the transpyloric plane, and that of the inferior mesenteric artery lies at the level of the body of the third lumbar vertebra, in the subcostal plane.
Renal arteries
The renal arteries can be projected as broad lines running laterally for 4 cm from the aorta just inferior to the transpyloric plane; the left inclines across the plane.
Iliac arteries
The surface projection of the common iliac artery corresponds to the superior one-third of a broad line, which is laterally slightly convex, from the aortic bifurcation to a point mid-way between the anterior superior iliac spine and the pubic symphysis. The external iliac artery corresponds to the inferior two-thirds of this line.
Inferior vena cava
The inferior vena cava starts at the level of the body of the fifth lumbar vertebra, usually in the transtubercular plane (2.5 cm below the supracristal plane). From this level it can be represented by a band, running vertically and 2.5 cm to the right of the midline. The inferior vena cava leaves the abdomen by traversing the diaphragm at the level of the eighth thoracic vertebra, directly behind the sternal extremity of the right sixth costal cartilage.
CLINICAL PROCEDURES
Pneumoperitoneum for laparoscopy
The establishment of a pneumoperitoneum is frequently performed by accessing the peritoneal cavity at the level of the umbilicus. Most commonly done just below the umbilicus, the incision through the linea alba allows access to the peritoneum at a point where there is relatively little extraperitoneal fat present and where the linea alba is relatively wide.
Surgical incisions
Most surgical incisions are performed according to surgical imperatives rather than anatomical constraints. The paramedian incision has been used to decrease the risk of incisional hernia. The skin and anterior fascial incision is made over the belly of rectus abdominis, at the level required. The muscle is displaced laterally, in order not to interfere with its neurovascular supply (which enters laterally), and the posterior rectus sheath is then incised in the paramedian plane. This approach allows rectus abdominis to lie between the two fascial incisions once the wound is closed.
Intestinal stomas (ileostomy, colostomy, caecostomy)
Where possible, intestinal stomas are usually formed through trans-rectus incisions. A cruciate incision is made in the anterior rectus sheath and the muscle fibres parted until the posterior sheath is reached. The latter is then incised without injury to the epigastric vessels (inferior or superior, depending on the level). This incision offers the advantage that fibres of rectus abdominis act to support the stoma: they provide a dynamic, contractile surround that tends to reduce the risk of herniation occurring around the stoma.
Suprapubic catheterization
The urinary bladder is commonly accessed for short- or long-term catheterization through the anterior abdominal wall. As the bladder fills, the upper part of its dome comes to lie in the preperitoneal space in the suprapubic region, and it can be relatively easily accessed via transcutaneous puncture in the midline (because there are no major neurovascular structures at this level and the linea alba is relatively thin).