STRUCTURE AND FUNCTION OF THE PULMONARY SYSTEM


The pulmonary system consists of upper and lower airways, the chest wall, and pulmonary circulation. In addition, the respiratory center of the central nervous system and the phrenic nerve participate in pulmonary function by providing the neurochemical control of breathing. The primary function of the pulmonary system is the exchange of gases between the environmental air and the blood. There are three steps in this process: (1) ventilation, the movement of air into and out of the lungs; (2) diffusion, the movement of gases between air spaces in the lungs and the bloodstream; and (3) perfusion, the movement of blood into and out of the capillary beds of the lungs to body organs and tissues. The first two functions are carried out by the pulmonary system and the third by the cardiovascular system (see Chapter 29). Normally the pulmonary system functions efficiently under a variety of conditions and with little energy expenditure.
STRUCTURES OF THE PULMONARY SYSTEM
The pulmonary system is made up of the upper airways, two lungs, the lower airways, and the blood vessels that serve them (Figure 32-1), and the chest wall, or thoracic cage. The lungs are divided into lobes: three in the right lung (upper, middle, lower) and two in the left lung (upper, lower). Each lobe is further divided into segments and lobules. The space between the lungs, which contains the heart, great vessels, and esophagus, is called the mediastinum. A set of conducting airways, called bronchi, deliver air to each section of the lung. The lung tissue that surrounds the airways supports them, preventing their distortion or collapse as gas moves in and out during ventilation.
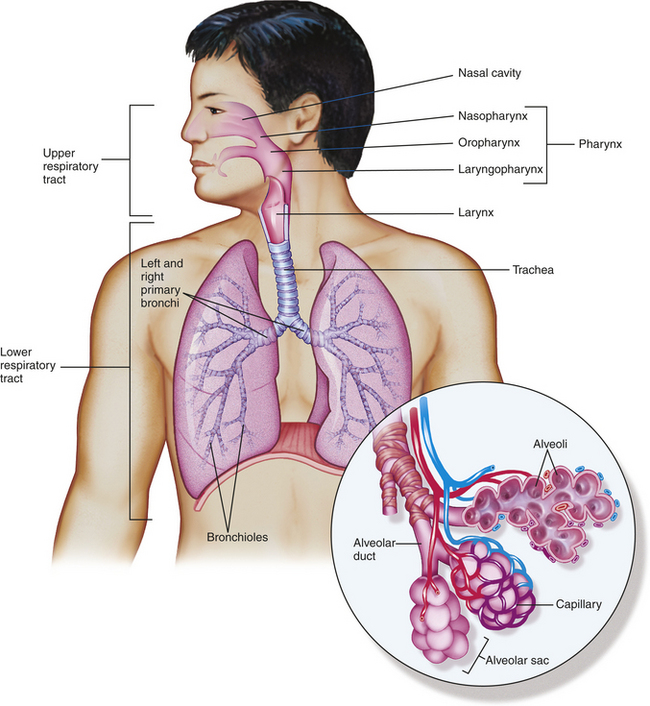
Figure 32-1 Structural plan of the respiratory system. Inset shows alveolar sacs where the interchange of oxygen and carbon dioxide takes place through the walls of the grapelike alveoli. Capillaries surround the alveoli. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
The lungs are protected from a variety of exogenous contaminants by a series of mechanical barriers (Table 32-1). These defense mechanisms are so effective that in the healthy individual, contamination of the lung tissue itself is unusual. (Other mechanisms of self-defense are discussed in Chapters 6 and 7.)
Table 32-1
| Structure or Substance | Mechanism of Defense |
| Upper respiratory tract mucosa | Maintains constant temperature and humidification of gas entering the lungs; traps and removes foreign particles, some bacteria, and noxious gases from inspired air |
| Nasal hairs and turbinates | Trap and remove foreign particles, some bacteria, and noxious gases from inspired air |
| Mucous blanket | Protects trachea and bronchi from injury; traps most foreign particles and bacteria that reach the lower airways |
| Cilia | Propel mucous blanket and entrapped particles toward the oropharynx, where they can be swallowed or expectorated |
| Alveolar macrophages | Ingest and remove bacteria and other foreign material from alveoli by phagocytosis (see Chapter 6) |
| Irritant receptors in nares (nostrils) | Stimulation by chemical or mechanical irritants triggers sneeze reflex, which results in rapid removal of irritants from nasal passages |
| Irritant receptors in trachea and large airways | Stimulation by chemical or mechanical irritants triggers cough reflex, which results in removal of irritants from the trachea and large airways |
Conducting Airways
The conducting airways are the portion of the pulmonary system that provides a passage for the movement of air into and out of the gas-exchange portions of the lung. They consist of upper and lower airways. The nasopharynx, oropharynx, and related structures often are called the upper airway (Figure 32-2). These structures are lined with a ciliated mucosa with a very rich vascular supply. The mucosal lining warms and humidifies inspired air and removes foreign particles from it as it passes into the lungs. During quiet breathing, gas usually flows through the nose, nasopharynx, and oropharynx to the lower airways. The mouth and oropharynx provide for ventilation when the nose is obstructed or when increased flow is required, such as during exercise. Filtering and humidifying are not, however, as efficient with mouth breathing.
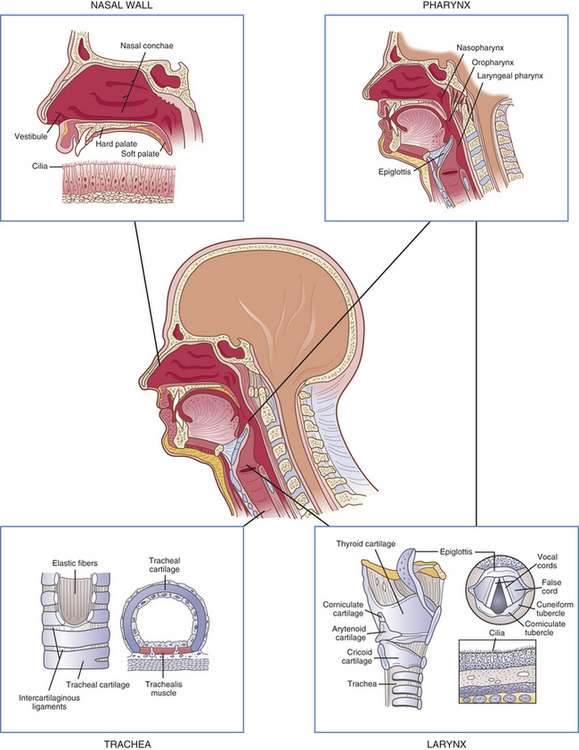
Figure 32-2 Structures of the upper airway. (Redrawn from Thompson JM et al: Mosby’s clinical nursing, ed 5, St Louis, 2002, Mosby.)
The larynx connects the upper and lower airways. The structure of the larynx consists of the endolarynx and its surrounding triangular-shaped bony and cartilaginous structures. The endolarynx is formed by two pairs of folds that form the false vocal cords (supraglottis) and the true vocal cords. The slit-shaped space between the true cords forms the glottis (see Figure 32-2). The vestibule is the space above the false vocal cords. The laryngeal box is formed of three large cartilages—the epiglottis, thyroid, and cricoid—and three smaller cartilages—the arytenoid, corniculate, and cuneiform—that are connected by ligaments. The supporting cartilages prevent collapse of the larynx during inspiration and swallowing. The internal laryngeal muscles control vocal cord length and tension, and the external laryngeal muscles move the larynx as a whole. Both sets of muscles are important to swallowing, respiration, and vocalization.1 The internal muscles contract during swallowing to prevent aspiration into the trachea and contribute to voice pitch.
The trachea, which is supported by U-shaped cartilage, connects the larynx to the bronchi, the conducting airways of the lungs. The trachea divides into the two main airways, or bronchi, at the carina (see Figure 32-1). This area is very sensitive and when stimulated can cause coughing and airway narrowing. The right main bronchus extends from the trachea more vertically than the left main bronchus, so that aspirated fluids or foreign particles tend to enter the right lung rather than the left. The right and left main bronchi enter the lungs at the hila, or “roots” of the lungs, along with the pulmonary blood and lymphatic vessels. From the hila the main bronchi branch into lobar bronchi, then to segmental and subsegmental bronchi, and finally end at the sixteenth division in the smallest of the conducting airways, the terminal bronchioles (Figure 32-3). With these multiple divisions, the cross-sectional area of the airways increases to 20 times that of the trachea. This results in decreased velocity or airflow into the gas-exchange portion of the lung and allows for optimal gas diffusion.2
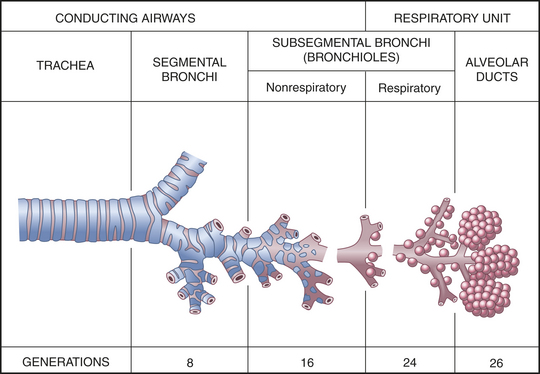
Figure 32-3 Structures of the lower airway. (Redrawn from Thompson JM et al: Mosby’s clinical nursing, ed 5, St Louis, 2002, Mosby.)
The bronchial walls have three layers: an epithelial lining, a smooth muscle layer, and a connective tissue layer. In the large bronchi (to approximately the tenth division), the connective tissue layer contains cartilage. The epithelial lining of the bronchi contains single-celled exocrine glands—the mucus-secreting goblet cells—and ciliated cells. High columnar pseudostratified epithelium lines the larger airways, changing to columnar cuboidal epithelium in the bronchioles (types of epithelia are illustrated in Chapter 1). The submucosal glands of the bronchial lining also produce mucus, contributing to the mucous blanket that covers the bronchial epithelium. The ciliated epithelial cells rhythmically beat this mucous blanket toward the trachea and pharynx, where it can be swallowed or expectorated by coughing. Foreign particles and microorganisms that are not expelled by mucociliary clearance and coughing are attacked by cellular components of the inflammatory response and antibodies of the secretory immune system (see Chapter 6). The biochemical mediators released early in inflammation also play a part in antibody-mediated hypersensitivity reactions, such as asthma, because they stimulate bronchial smooth muscles to constrict.
With branching, the layers of epithelium that line the bronchi become thinner (Figure 32-4). Ciliated cells and goblet cells become more sparse, and smooth muscle and connective tissue layers thin toward the terminal bronchioles.2
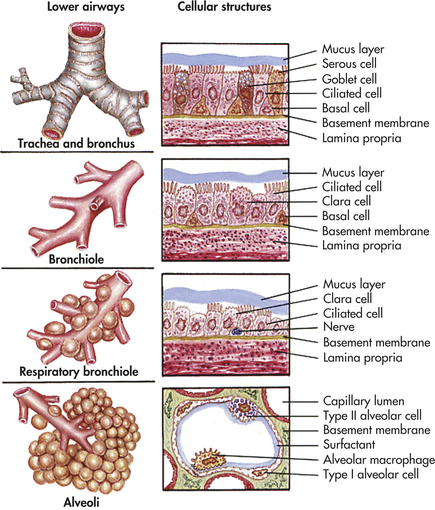
Gas-Exchange Airways
The conducting airways terminate in gas-exchange airways, where oxygen (O2) enters the blood and carbon dioxide (CO2) is removed from it. The gas-exchange airways are made up of respiratory bronchioles, alveolar ducts, and alveoli. These structures together are sometimes called the acinus (see Figure 32-3), and all of them participate in gas exchange.3
The bronchioles from the sixteenth through the twenty-third divisions contain increasing numbers of alveoli and are called respiratory bronchioles. The walls of the respiratory bronchioles are very thin, consisting of an epithelial layer devoid of cilia and goblet cells, very little smooth muscle fiber, and a very thin and elastic connective tissue layer. These bronchioles end in alveolar ducts, which lead to alveolar sacs made up of numerous alveoli.
The alveoli are the primary gas-exchange units of the lung, where oxygen enters the blood and CO2 is removed (Figure 32-5). Tiny passages called pores of Kohn permit some air to pass through the septa from alveolus to alveolus, promoting collateral ventilation and even distribution of air among the alveoli. The lungs contain approximately 25 million alveoli at birth and 300 million by adulthood.
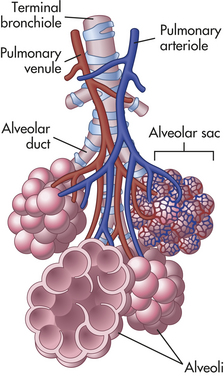
Figure 32-5 Alveoli. Bronchioles subdivide to form tiny tubes called alveolar ducts that end in clusters of alveoli called alveolar sacs. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
The alveolar septa consist of an epithelial layer and a thin, elastic basement membrane but no muscle layer (Figure 32-6). Two major types of epithelial cells appear in the alveolus. Type I alveolar cells provide structure, and type II alveolar cells secrete surfactant, a lipoprotein that coats the inner surface of the alveolus and facilitates its expansion during inspiration, lowers alveolar surface tension at end-expiration, and, thereby, prevents lung collapse.1–4
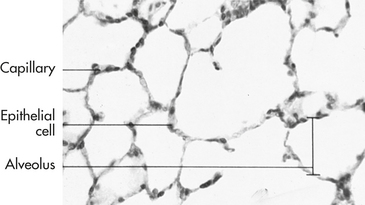
Figure 32-6 Photomicrograph of lung, showing several alveoli. Note the proximity of the capillary to the alveolar wall. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
Like the bronchi, alveoli contain cellular components of inflammation and immunity, particularly the mononuclear phagocytes. The mononuclear phagocytes of the lungs are called alveolar macrophages. These cells ingest foreign material that reaches the alveolus and prepare it for removal through the lymphatics.1 (Phagocytosis and the mononuclear phagocyte system are described in Chapters 6 and 7.)
Pulmonary and Bronchial Circulation
The pulmonary circulation facilitates gas exchange, delivers nutrients to lung tissues, acts as a reservoir for the left ventricle, and serves as a filtering system that removes clots, air, and other debris from the circulation (Figure 32-7).
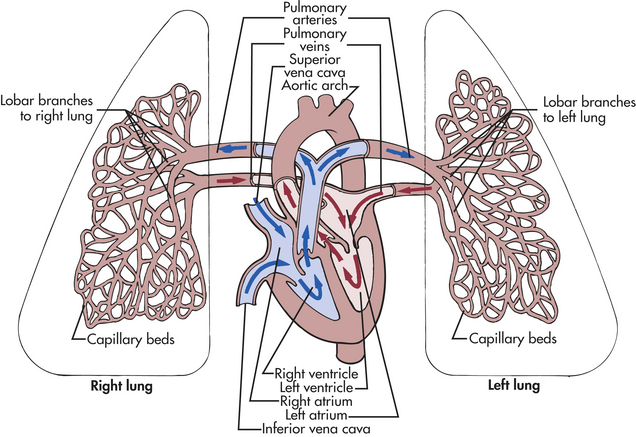
Figure 32-7 The pulmonary circulation. The right and left pulmonary veins and arteries and the branching capillaries are illustrated. Note the pulmonary artery carries venous blood, and the pulmonary vein carries arterial blood.
Although the entire cardiac output from the right ventricle goes into the lungs, the pulmonary circulation has a lower pressure and resistance than the systemic circulation. Pulmonary arteries are exposed to about one fifth the pressure of the systemic circulation and have a much thinner muscle layer. (Systemic vessels are described in Chapter 29.) Mean pulmonary artery pressure is 18 mmHg; mean aortic pressure is 90 mmHg. About one third of the pulmonary vessels are filled with blood (perfused) at any given time. More vessels become perfused when right ventricular cardiac output increases. Therefore, increased delivery of blood to the lungs does not normally increase mean pulmonary artery pressure.
The pulmonary artery divides and enters the lung at the hilus with each main bronchus and branches with the bronchus at every division so that every bronchus and bronchiole has an accompanying artery or arteriole. The arterioles, less than 1 mm in diameter, regulate blood flow through their respective capillary beds.
The arterioles divide at the terminal bronchiole to form a network of pulmonary capillaries around the acinus. The capillaries are an integral part of the alveolar septa. Capillary walls consist of an endothelial layer and a thin basement membrane, which often fuses with the basement membrane of the alveolar septum (see Figure 32-6). This results in very little separation between blood in the capillary and gas in the alveolus.
The shared alveolar and capillary walls compose the alveolocapillary membrane, a very thin membrane made up of the alveolar epithelium, the alveolar basement membrane, an interstitial space, the capillary basement membrane, and the capillary endothelium (Figure 32-8). These extremely thin alveolar walls are easily damaged and can leak plasma and blood into the alveolar space. Gas exchange occurs across the alveolocapillary membrane. With normal perfusion approximately 100 ml of blood in the pulmonary capillary bed is spread very thinly over 70 to 100 m2 of alveolar surface area. The alveolocapillary membrane efficiently exposes large quantities of blood to gas in the alveoli. Any disorder that thickens the membrane impairs gas exchange.
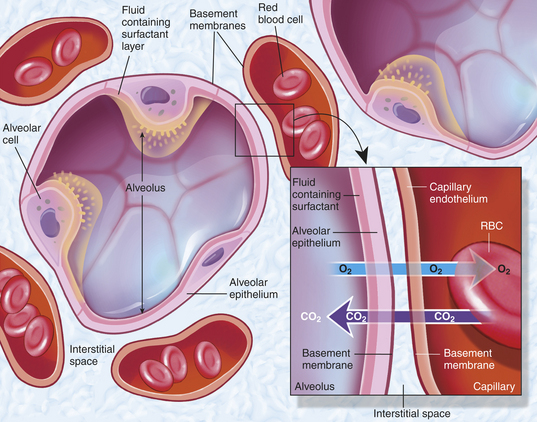
Figure 32-8 Section through the alveolar septum (gas-exchange membrane). Inset shows a magnified view of the respiratory membrane composed of the alveolar wall (fluid coating, epithelial cells, basement membrane), interstitial fluid, and wall of a pulmonary capillary (basement membrane, endothelial cells). The gases—carbon dioxide (CO2) and oxygen (O2)—diffuse across the respiratory membrane. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
Each pulmonary vein drains several pulmonary capillaries. Unlike the pulmonary arteries, which follow the branching bronchi, pulmonary veins are dispersed randomly throughout the lung and then leave the lung at the hila and enter the left atrium. They are similar to veins in the systemic circulation, but they have no valves.
The bronchial circulation is part of the systemic circulation. It supplies nutrients to the conducting airways, nerves, lymph nodes, large pulmonary vessels, and membranes (pleurae) that surround the lungs.2 The bronchial circulation is unique in that not all of its capillaries drain into its own venous system. Some of the bronchial capillaries empty into the pulmonary vein and contribute to the normal venous admixture (mixing of oxygenated and deoxygenated blood) or right-to-left shunt (right-to-left shunts are described in Chapter 33). The bronchial circulation does not participate in gas exchange.
Lung vasculature also includes deep and superficial lymphatic capillaries. The deep lymphatic capillaries begin at the level of the terminal bronchioles; there are no lymphatic structures in the acinus. Fluid and alveolar macrophages migrate from the alveoli to the terminal bronchioles, where they enter the lymphatic system. The superficial lymphatic capillaries drain the membrane that surrounds the lungs. Both deep and superficial lymphatic vessels leave the lung at the hilus. The lymphatic system plays an important role in keeping the lung free of fluid. (The lymphatic system is described in Chapter 29.)
Chest Wall and Pleura
The chest wall (skin, ribs, intercostal muscles) protects the lungs from injury, and its muscles, in conjunction with the diaphragm, perform the muscular work of breathing. The thoracic cavity is contained by the chest wall and encases the lungs (Figure 32-9). A serous membrane called the pleura adheres firmly to the lungs. It then folds over itself and attaches firmly to the chest wall. The membrane covering the lungs is the visceral pleura; that lining the thoracic cavity is the parietal pleura. The area between the two pleurae is called the pleural space, or pleural cavity. Normally only a thin layer of fluid secreted by the pleura (pleural fluid) fills the pleural space. This lubricates the pleural surfaces, allowing the two layers to slide over each other without separating. Pressure in the pleural space is usually negative or subatmospheric (−4 to −10 mmHg).
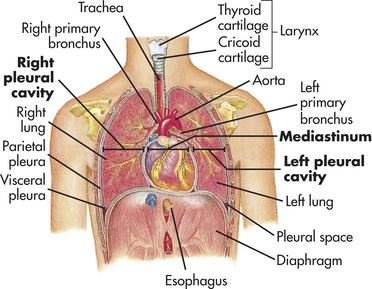
Figure 32-9 Thoracic (chest) cavity and related structures. The thoracic cavity is divided into three subdivisions (left and right pleural divisions and mediastinum) by a partition formed by a serous membrane called the pleura. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 3, St Louis, 1996, Mosby.)
FUNCTIONS OF THE PULMONARY SYSTEM
The pulmonary system functions to (1) ventilate the alveoli, (2) diffuse gases into and out of the blood, and (3) perfuse the lungs so that the organs and tissues of the body receive blood that is rich in oxygen and low in CO2. Each component of the pulmonary system contributes to one or more of these functions (Figure 32-10).
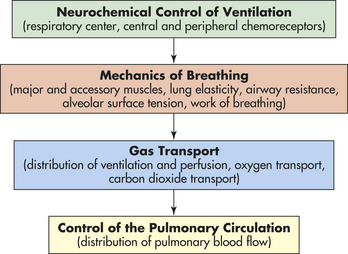
Figure 32-10 Functional components of the respiratory system. The central nervous system responds to neurochemical stimulation of ventilation and sends signals to the chest wall musculature. The response of the respiratory system to these impulses is influenced by several factors that affect the mechanisms of breathing and therefore affect the adequacy of ventilation. Gas transport between the alveoli and pulmonary capillary blood depends on a variety of physical and chemical activities. The control of the pulmonary circulation plays a role in the appropriate distribution of blood flow.
Ventilation
Ventilation is the mechanical movement of gas or air into and out of the lungs. Ventilation often is misnamed respiration, which is actually the exchange of O2 and CO2 during cellular metabolism. “Respiratory rate” is actually the ventilatory rate, or the number of times gas is inspired and expired per minute. The amount of effective ventilation is calculated by multiplying the ventilatory rate (breaths per minute) by the volume of air per breath (liters per breath, tidal volume). This is called the minute volume or minute ventilation and is expressed in liters per minute.
CO2, the gaseous form of carbonic acid (H2CO3), is a product of cellular metabolism. The lung eliminates about 10,000 milliequivalents (mEq) of carbonic acid per day in the form of CO2, which is produced at the rate of approximately 200 ml/minute. CO2 elimination is necessary to maintain a normal arterial CO2 (PaCO2) of 40 mmHg and normal acid-base balance (see Chapter 3 for a discussion of acid-base regulation).
The adequacy of alveolar ventilation cannot be accurately determined by observation of ventilatory rate, pattern, or effort. If a healthcare professional needs to determine the adequacy of ventilation, an arterial blood gas analysis must be performed to measure PaCO2.
Neurochemical Control of Ventilation
The mechanisms that control respiration are very complex.1–5 Breathing is usually involuntary because homeostatic changes in the ventilatory rate and volume are adjusted automatically by the nervous system to maintain normal gas exchange. Voluntary breathing is necessary for talking, singing, laughing, and holding one’s breath.
The respiratory center in the brainstem controls respiration by transmitting impulses to the respiratory muscles, causing them to contract and relax (Figure 32-11). The respiratory center is composed of several groups of neurons located bilaterally in the brainstem: the dorsal respiratory group (DRG), the ventral respiratory group (VRG), the pneumotaxic center, and the apneustic center.1,4 The basic automatic rhythm of respiration is set by the DRG, a cluster of inspiratory nerve cells located in the medulla that sends efferent impulses to the diaphragm and inspiratory intercostal muscles. The DRG also receives afferent impulses from peripheral chemoreceptors in the carotid and aortic bodies, which detect the PaCO2 and the amount of oxygen in the arterial blood (PaO2). In addition, several different types of receptors in the lungs stimulate the VRG through afferent nerves. The VRG, also located in the medulla, contains inspiratory and expiratory neurons. It is almost inactive during normal, quiet respiration, becoming active when increased ventilatory effort is required. The pneumotaxic center and apneustic center, situated in the pons, do not generate primary rhythm but rather act as modifiers of the inspiratory depth and rate established by the medullary centers.4 Breathing can be modified by input from the cortex, the limbic system, and the hypothalamus, and the pattern of breathing can be influenced by emotion and by disease.
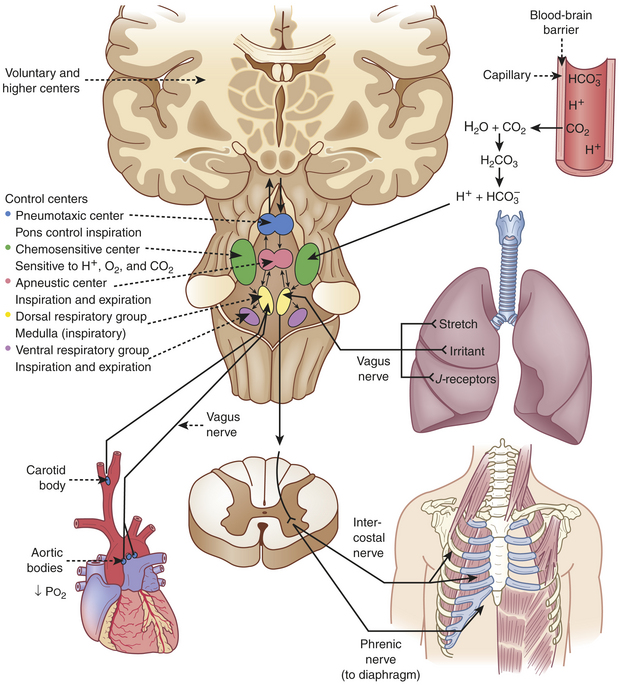
Lung Receptors: Three types of lung receptors send impulses from the lungs to the dorsal respiratory group:
1. Irritant receptors are found in the epithelium of the conducting airways. They are sensitive to noxious aerosols (vapors), gases, and particulate matter (e.g., inhaled dusts), which cause them to initiate the cough reflex. When stimulated, irritant receptors also cause bronchoconstriction and increased ventilatory rate. These receptors are located primarily in the proximal larger airways and are nearly absent in the distal airways; thus it is possible for secretions to accumulate in the distal respiratory tree without initiating cough.
2. Stretch receptors are located in the smooth muscles of airways and are sensitive to increases in the size or volume of the lungs. They decrease ventilatory rate and volume when stimulated, an occurrence sometimes referred to as the Hering-Breuer expiratory reflex. This reflex is active in newborns and assists with ventilation. In adults, this reflex is active only at high tidal volumes (such as with exercise and mechanical ventilation) and may play a role in protecting against excess lung inflation.6 Stretch receptors called rapidly adapting receptors have been found to be an important mediator of cough.7
3. J-receptors (juxtapulmonary capillary receptors) are located near the capillaries in the alveolar septa. They are sensitive to increased pulmonary capillary pressure, which stimulates them to initiate rapid, shallow breathing; hypotension; and bradycardia.4
The lung is innervated by the autonomic nervous system (ANS). Fibers of the sympathetic division of the ANS in the lung branch from the upper thoracic and cervical ganglia of the spinal cord. Fibers of the parasympathetic division of the ANS travel in the vagus nerve to the lung. (Structures and function of the ANS are discussed in detail in Chapter 14.) The parasympathetic and sympathetic divisions of the ANS control airway caliber (interior diameter of the airway lumen) by stimulating bronchial smooth muscle to contract or relax. The parasympathetic receptors cause smooth muscle to contract, whereas sympathetic receptors cause it to relax. Bronchial smooth muscle tone depends on equilibrium, that is, equal stimulation of contraction and relaxation. The parasympathetic division of the ANS is the main controller of airway caliber under normal conditions.4 Constriction occurs if the irritant receptors in the airway epithelium are stimulated by irritants in inspired air, by endogenous substances (e.g., histamine, serotonin, prostaglandins), by many drugs, and by humoral substances.
Chemoreceptors: Chemoreceptors monitor pH, PaCO2, and PaO2Central chemoreceptors monitor arterial blood indirectly by sensing changes in the pH of cerebrospinal fluid (CSF).5 They are located near the respiratory center and are sensitive to hydrogen ion concentration in the CSF. (Chapter 3 describes the relationship between ions and the pH, or acid-base status, of body fluids.) The pH, or concentration of hydrogen ions in the CSF, reflects PaCO2 because, unlike H+ ions, CO2 in arterial blood diffuses across the blood-brain barrier (the capillary wall separating blood from cells of the central nervous system) into the CSF until the partial pressure of CO2 (PCO2) is equal on both sides. CO2 that has entered the CSF combines with H2O to form carbonic acid, which subsequently dissociates into hydrogen ions that are capable of stimulating the central chemoreceptors. In this way PaCO2 regulates ventilation through its effect on the pH (hydrogen ion content) of the CSF.1,4
If alveolar ventilation is inadequate, PaCO2 increases. CO2 diffuses across the blood-brain barrier until PCO2 in the blood and CSF reaches equilibrium. As the central chemoreceptors sense the resulting decrease in pH (increase in hydrogen ion concentration), they stimulate the respiratory center to increase the depth and rate of ventilation. Increased ventilation causes the PaCO2 to decrease below that of the CSF, and CO2 diffuses back out of the CSF, returning its pH to normal.
The central chemoreceptors are sensitive to very small changes in the pH of CSF (equivalent to a 1- to 2-mmHg change in PCO2) and are able to maintain a normal PaCO2 under many different conditions, including strenuous exercise. If inadequate ventilation, or hypoventilation, is long term (e.g., in chronic obstructive pulmonary disease), these receptors become insensitive to small changes in PaCO2 and regulate ventilation poorly. In addition, prolonged increases in PaCO2 result in renal compensation through bicarbonate retention. This bicarbonate gradually diffuses into the CSF, where it normalizes the pH and negates the effect on ventilatory drive.1,4
The peripheral chemoreceptors are located in aortic bodies, the aortic arch, and carotid bodies at the bifurcation of the carotids, near the baroreceptors (see Chapter 29). Although the peripheral chemoreceptors are sensitive to changes in PaCO2 and pH, they are primarily sensitive to oxygen levels in arterial blood (PaO2) and are responsible for all of the increase in ventilation that occurs in response to arterial hypoxemia.4 As PaO2 and pH decrease, peripheral chemoreceptors, particularly in the carotid bodies, send signals to the respiratory center to increase ventilation. The peripheral chemoreceptors are not as sensitive as the central chemoreceptors. The PaO2 must drop well below normal (to approximately 60 mmHg) before the peripheral chemoreceptors have much influence on ventilation. If PaCO2 is elevated as well, however, ventilation increases much more than it would in response to either abnormality alone. The peripheral chemoreceptors become the major stimulus to ventilation when the central chemoreceptors are “reset” by chronic hypoventilation.
Mechanics of Breathing
The mechanical aspects of inspiration and expiration are known collectively as the mechanics of breathing and involve (1) major and accessory muscles of inspiration and expiration, (2) elastic properties of the lungs and chest wall, and (3) resistance to airflow through the conducting airways. Alterations in any of these properties increase the work of breathing, or the metabolic energy that must be exerted to achieve adequate ventilation and oxygenation of the blood.
Major and Accessory Muscles: The major muscles of inspiration are the diaphragm and the external intercostal muscles (muscles between the ribs) (Figure 32-12). The diaphragm is a dome-shaped muscle that separates the abdominal and thoracic cavities. When the diaphragm contracts, it flattens downward, increasing the volume of the thoracic cavity, and creates a negative pressure that draws gas into the lungs through the upper airways and trachea. Contraction of external intercostal muscles elevates the anterior portion of the ribs. This increases the volume of the thoracic cavity by increasing its front-to-back (anteroposterior [AP]) diameter. Although the external intercostal muscles may contract during quiet breathing, inspiration at rest usually is assisted by the diaphragm only.
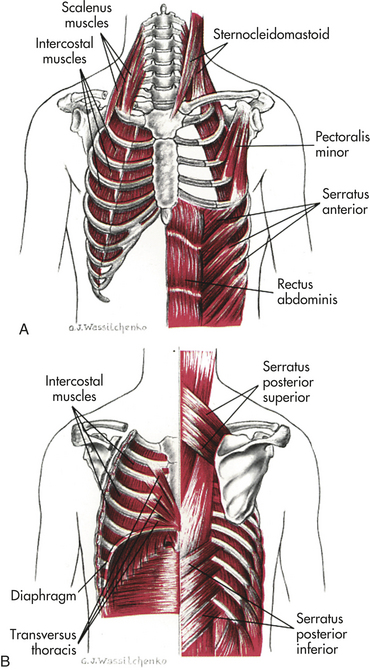
Figure 32-12 Muscles of ventilation. A, Anterior view. B, Posterior view. (From Wilson SF, Thompson JM: Respiratory disorders, St Louis, 1990, Mosby.)
The accessory muscles of inspiration are the sternocleidomastoid and scalene muscles. Like the external intercostal muscles, these muscles enlarge the thorax by increasing its AP diameter. The accessory muscles of inspiration assist inspiration when minute volume (volume of air inspired and expired per minute) is very high, such as during strenuous exercise or when the work of breathing is increased because of disease. The accessory muscles do not increase the volume of the thorax as efficiently as the diaphragm does.
There are no major muscles of expiration because normal, relaxed expiration is passive and requires no muscular effort. The accessory muscles of expiration, the abdominal and internal intercostal muscles, assist expiration when minute volume is high, during coughing, or when airway obstruction is present. When the abdominal muscles contract, intra-abdominal pressure increases, pushing up the diaphragm and decreasing the volume of the thorax. The internal intercostal muscles pull down the anterior ribs, decreasing the AP diameter of the thorax.
Alveolar Surface Tension: Surface tension occurs at any gas-liquid interface and refers to the tendency for liquid molecules that are exposed to air to adhere to one another. This phenomenon can be seen, for example, in a glass of liquid that is about to overflow or in the way liquids “bead” when splashed on a waterproof surface. In both examples this phenomenon decreases the surface area exposed to the air.
Within a sphere, such as an alveolus, surface tension tends to make expansion difficult. According to the law of Laplace, the pressure (P) required to inflate a sphere is equal to two times the surface tension (2T) divided by the radius (r) of the sphere, or P = (2T/r).1 As the radius of the sphere (or alveolus) becomes smaller, more and more pressure is required to inflate it. If the alveoli were lined with a water-like fluid, taking breaths would be extremely difficult.
Alveolar ventilation, or distention, is made possible by surfactant, which lowers the surface tension by coating the air-liquid interface in the alveoli. Surfactant, a lipoprotein produced by type II alveolar cells (see Figure 32-4), has a detergent-like effect that separates the liquid molecules, thereby decreasing alveolar surface tension.
Surfactant lines the alveolar side of the alveolocapillary membrane and, in effect, reverses Laplace’s law. As the radius of a surfactant-lined sphere (alveolus) grows smaller, the surface tension decreases, and as the radius grows larger, the surface tension increases. This occurs because the surfactant molecules have much weaker intermolecular attraction compared with the liquid molecules. The surfactant molecules occupy most of the air-fluid interface and disrupt the intermolecular forces that tend to collapse the alveoli. Therefore, the alveoli are much easier to inflate at low lung volumes (i.e., after expiration) than at high volumes (i.e., after inspiration). If surfactant production is disrupted or surfactant is not
produced in adequate quantities, alveolar surface tension increases and results in alveolar collapse, decreased lung expansion, increased work of breathing, and severe gas-exchange abnormalities (see What’s New? Update on Surfactant).
The decrease in surface tension caused by surfactant is also responsible for keeping the alveoli free of fluid. In the absence of surfactant, the surface tension tends to attract fluid into the alveoli. In addition, surfactant participates in host defense against respiratory pathogens.1,8
Elastic Properties of the Lung and Chest Wall: The lung and chest wall have elastic properties that permit expansion during inspiration and return to resting volume during expiration. The elasticity of the lungs is caused both by elastin fibers in the alveolar walls and surrounding the small airways and pulmonary capillaries and by surface tension at the alveolar air-liquid interface.1 The elasticity of the chest wall is the result of the configuration of its bones and musculature.
Elastic recoil is the tendency of the lungs to return to the resting state after inspiration. Normal elastic recoil permits passive expiration, eliminating the need for major muscles of expiration. Passive elastic recoil may be insufficient during labored breathing (high minute volume), in which case the accessory muscles of expiration may be needed. The accessory muscles also are used if disease comprises elastic recoil (e.g., in emphysema) or blocks the conducting airways.
Normal elastic recoil depends on an equilibrium between opposing forces of recoil in the lungs and chest wall. Under normal conditions the chest wall tends to recoil by expanding outward. This can be observed readily during open heart surgery. When the sternum is split to open the thoracic cavity, the chest wall moves outward laterally. The tendency of the chest wall to recoil by expanding is balanced by the tendency of the lungs to recoil or collapse around the hila. This reaction is caused by elastic recoil and surface tension in the alveoli. The tendency of the lungs to collapse can be demonstrated if the chest is opened without mechanically ventilating the lungs (e.g., at postmortem examination). As the thorax is opened, the lungs immediately collapse, like inflated balloons that have been released. The opposing forces of the chest wall and lungs create, in part, the small negative intrapleural pressure.
Balance between the outward recoil of the chest wall and inward recoil of the lungs occurs at the resting level, at the end of expiration. During inspiration the diaphragm and intercostal muscles contract, air flows into the lungs, and the chest wall expands. Muscular effort is needed to overcome the resistance of the lungs to expansion. During expiration the muscles relax and the elastic recoil of the lungs causes the thorax to decrease in volume until, once again, balance between the chest wall and lung recoil forces is reached1 (Figure 32-13).
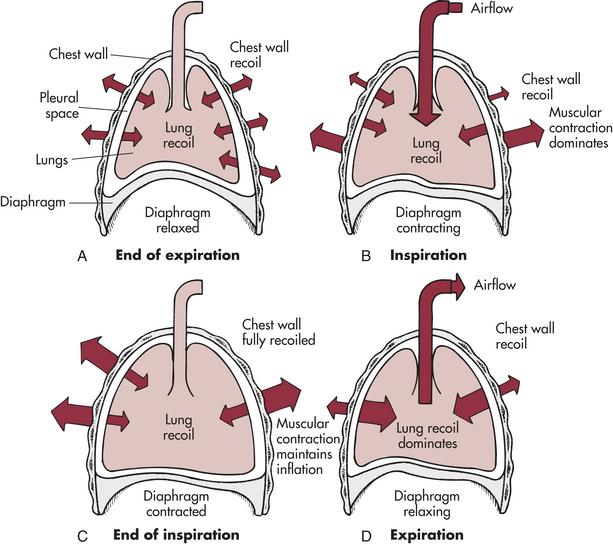
Figure 32-13 Interaction of forces during inspiration and expiration. A, Outward recoil of the chest wall equals inward recoil of the lungs at the end of expiration. B, During inspiration, contraction of respiratory muscles, assisted by chest wall recoil, overcomes tendency of lungs to recoil. C, At the end of inspiration, respiratory muscle contraction maintains lung expansion. D, During expiration, respiratory muscles relax, allowing elastic recoil of the lungs to deflate the lungs.
Compliance is the measure of lung and chest wall distensibility. It represents the relative ease with which these structures can be stretched. Compliance is therefore the reciprocal of elasticity. Compliance is determined by alveolar surface tension and the elastic recoil of the lung and chest wall. It can be measured with the following formula:
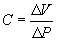
where C = compliance in liters per centimeter of water, ΔV = volume change (usually tidal volume), and ΔP = pressure change (airway or pleural pressure) in centimeters of water.1,9
Increased compliance indicates that the lungs or chest wall is abnormally easy to inflate and has lost some elastic recoil. A decrease indicates that the lungs or chest wall is abnormally stiff or difficult to inflate. Compliance is increased in emphysema and decreased in acute respiratory distress syndrome, pneumonia, pulmonary edema, and fibrosis. (These disorders are described in Chapter 33.)
Airway Resistance: Airway resistance, which is similar to resistance to blood flow (described in Chapter 29), is determined by the length, radius, and cross-sectional area of the airways and density, viscosity, and velocity of the gas (Poiseuille’s law). Resistance is computed by dividing change in pressure (P) by rate of flow (F), or R = {P/F} (Ohm’s law) and can easily be measured in the pulmonary function laboratory.5 Airway resistance is normally very low. One half to two thirds of total airway resistance occur in the nose. The next highest resistance is in the oropharynx and larynx. There is very little resistance in the conducting airways of the lungs because of their large cross-sectional area. Resistance increases as the diameter of the airways (total cross-sectional area) decreases; airway resistance increases when the diameter of the airways decreases. Bronchoconstriction, which increases airway resistance, can be caused by stimulation of parasympathetic receptors in the bronchial smooth muscle and by numerous irritants and inflammatory mediators.2 Bronchodilation, which decreases resistance to airflow, is caused by β2-adernergic receptor stimulation. Airway resistance also can be increased by edema of the bronchial mucosa and by airway obstructions such as mucus, tumors, or foreign bodies.
Work of Breathing: The work of breathing is determined by the muscular effort (and therefore oxygen and energy) required for ventilation. The work of breathing is normally very low but may increase considerably in disease states that disrupt the equilibrium between forces exerted by the lung and chest wall. More muscular effort is required when lung compliance is decreased (e.g., in pulmonary edema), chest wall compliance is decreased (e.g., in spinal deformity or obesity), or airways are obstructed by bronchospasm or mucous plugging (e.g., in asthma or bronchitis).9 An increase in the work of breathing can result in a marked increase in oxygen consumption and metabolic demand, which can cause significant morbidity in individuals with severe lung disease.
Measurement of Gas Pressure
A gas is made up of millions of molecules moving randomly. As they move, they collide with each other and the wall of the space in which they are contained. These collisions exert pressure. If more molecules are present in the space, the pressure, or number of collisions, increases (Figure 32-14). If the same number of gas molecules is contained in a small and a large container, the pressure is greater in the small container because more collisions occur in the smaller space. Heat increases the speed of the molecules, which increases the number of collisions. Therefore, pressure also increases at higher temperatures.
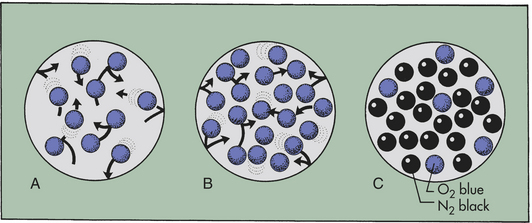
Figure 32-14 Relationship between number of gas molecules and pressure exerted by the gas in an enclosed space. (A), Theoretically 10 molecules of the same gas exert a total pressure of 10 within the space. (B), If the number of molecules is increased to 20, total pressure is 20. (C), If there are different gases in the space, each gas exerts a partial pressure: here the partial pressure of nitrogen (N2) is 18, that of oxygen (O2) is 6, and total pressure is 24.
Barometric pressure (PB) (atmospheric pressure) is the pressure exerted by gas molecules in air at specific altitudes. At sea level, barometric pressure is 760 mmHg. This number is the sum of the pressure exerted by each gas in the air at sea level. The portion of the total pressure exerted by any individual gas is its partial pressure (see Figure 32-14). At sea level the air is made up of oxygen (20.9%), nitrogen (78.1%), and a few other trace gases. The partial pressure of oxygen is equal to the percentage of oxygen in the air (20.9%) times the total pressure (760 mmHg), or 159 mmHg (760 × 0.209 = 158.84). (Symbols used in the measurement of gas pressures and pulmonary ventilation are defined in Table 32-2.)
Table 32-2
Common Pulmonary Abbreviations
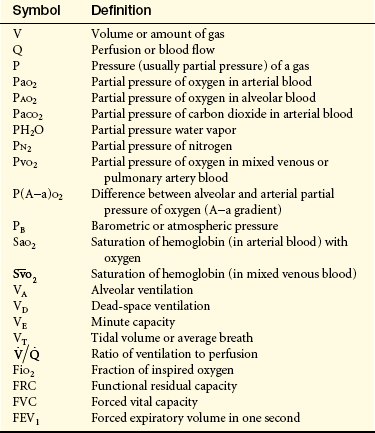
Subscripts identify the particular gas, volume, or pressure being discussed. A dot (•) means measurement over time, usually 1 minute.
The amount of water vapor contained in a gas mixture is determined by the temperature of the gas and is unrelated to barometric pressure. Gas that enters the lungs becomes saturated with water vapor (humidified) as it passes through the upper airway. At body temperature (37° C, 98.6° F), water vapor exerts a pressure of 47 mmHg. Because this is true regardless of total (barometric) pressure, the partial pressure of water vapor (always 47 mmHg) must be subtracted from the barometric pressure before the partial pressure of other gases in the mixture can be determined. In saturated air at sea level, the partial pressure of oxygen is therefore (760 − 47) × 0.209 = 149. All pressure and volume measurements made in pulmonary function laboratories specify the temperature and humidity of a gas at the time of measurement.
Many pressure measurements are stated as variations from barometric pressure, rather than percentages of it. On such scales, barometric pressure is considered zero, and pressure varies up or down from zero. Physiologic pressure measurements that involve fluids, rather than gases, are measured as variations from barometric pressure. For example, a systolic blood pressure of 120 mmHg indicates that systolic pressure is 120 mmHg above barometric pressure.
Gas Transport
Gas transport, the delivery of oxygen to the cells of the body and the removal of CO2, has four steps:
2. Diffusion of oxygen from the alveoli into the capillary blood
3. Perfusion of systemic capillaries with oxygenated blood
4. Diffusion of oxygen from systemic capillaries into the cells
Steps in the transport of CO2 occur in reverse order:
1. Diffusion of CO2 from the cells into the systemic capillaries
2. Perfusion of the pulmonary capillary bed by venous blood
If any step in gas transport is impaired by a respiratory or cardiovascular disorder, gas exchange at the cellular level is compromised.
Distribution of Ventilation and Perfusion
Effective gas exchange depends on an approximately even distribution of gas (ventilation) and blood (perfusion) in all portions of the lungs. The lungs are suspended from the hila in the thoracic cavity. When the individual is in an upright position (sitting or standing), gravity pulls the lungs down toward the diaphragm and compresses their lower portions or bases. The alveoli in the upper portions, or apices, of the lungs contain a greater residual volume of gas and are larger and less numerous than those in the lower portions. Because surface tension increases as the alveoli become larger, the larger alveoli in the upper portions of the lung are more difficult to inflate (less compliant) than the smaller alveoli in the lower portions of the lung. Therefore, during ventilation most of the tidal volume is distributed to the bases of the lungs, where compliance is greater.
The heart pumps against gravity to perfuse the pulmonary circulation. As blood is pumped into the lung apices of a sitting or standing individual, some blood pressure is dissipated in overcoming gravity. As a result, blood pressure at the apices is lower than that at the bases. Because greater pressure causes greater perfusion, the bases of the lungs are better perfused than the apices (Figure 32-15). Thus ventilation and perfusion are greatest in the same lung portions: the lower lobes. Ventilation and perfusion depend on body position. If a standing individual assumes a supine or side-lying position, the areas of the lungs that are then most dependent become the best ventilated and perfused.
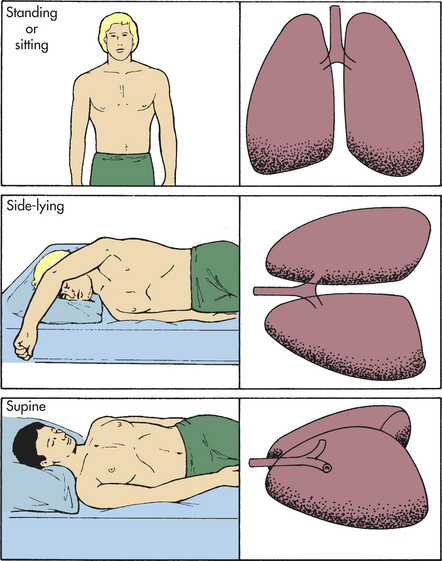
Figure 32-15 Pulmonary blood flow and gravity. The greatest volume of pulmonary blood flow will normally occur in the gravity-dependent areas of the lungs. Body position has a significant effect on the distribution of pulmonary blood flow.
Distribution of perfusion in the pulmonary circulation also is affected by alveolar pressure (gas pressure in the alveoli). The pulmonary capillary bed differs from the systemic capillary bed in that it is surrounded by gas-containing alveoli. If the gas pressure in the alveoli exceeds the blood pressure in the capillary, the capillary collapses and flow ceases. This is most likely to occur in portions of the lung where blood pressure is lowest and alveolar gas pressure is greatest, that is, the apex of the lung.
The lungs are divided into three zones on the basis of the relationships among all the factors affecting pulmonary blood flow. Alveolar pressure plus the forces of gravity, arterial blood pressure, and venous blood pressure affect the distribution of perfusion (Figure 32-16).
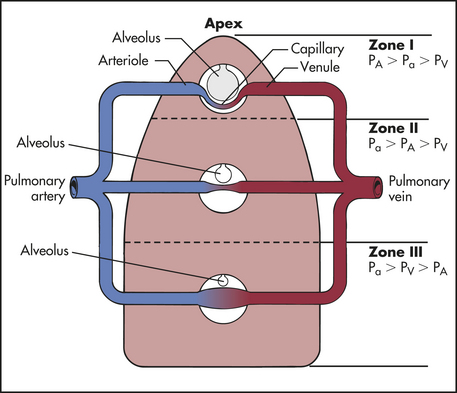
Figure 32-16 Gravity and alveolar pressure. Effects of gravity and alveolar pressure on pulmonary blood flow in the three lung zones. In zone I, alveolar pressure (PA) is greater than arterial and venous pressure, and no blood flow occurs. In zone II, arterial pressure (Pa) exceeds alveolar pressure, but alveolar pressure exceeds venous pressure (PV). Blood flow occurs in this zone, but alveolar pressure compresses the venules (venous ends of the capillaries). In zone III, both arterial and venous pressures are greater than alveolar pressure and blood flow fluctuates, depending on the difference between arterial and venous pressures.
Zone I is where alveolar pressure exceeds pulmonary arterial and venous pressures. The capillary bed collapses, and normal blood flow ceases. Normally zone I is a very small part of the lung at the apex. Zone II is the portion where alveolar pressure is greater than venous pressure but not greater than arterial pressure. Blood flows through zone II, but it is impeded to a certain extent by alveolar pressure. Zone II is normally above the level of the left atrium. In zone III arterial and venous pressures are greater than alveolar pressure and blood flow is not affected by alveolar pressure. Zone III is in the base of the lung. Blood flow through the pulmonary capillary bed increases in regular increments from the apex to the base.
Although blood flow and ventilation are greater at the base of the lungs than at the apices, they are not perfectly matched in any of the zones. Perfusion exceeds ventilation in the bases of the lungs, and ventilation exceeds perfusion in the apices of the lung. The relationship between ventilation and perfusion is expressed as a ratio called the ventilation-perfusion ratio, or 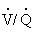 .1,4 The normal ratio is 0.8. This is the amount by which perfusion exceeds ventilation under normal conditions.
.1,4 The normal ratio is 0.8. This is the amount by which perfusion exceeds ventilation under normal conditions.
Oxygen Transport
Approximately 1000 ml (1 L) of oxygen is transported to the cells each minute. Oxygen is transported in the blood in two forms. A small amount dissolves in plasma, and the remainder binds to hemoglobin molecules. Without hemoglobin, oxygen would not reach the cells in amounts sufficient to maintain normal metabolic function. (Hemoglobin is discussed in detail in Chapter 25; cellular metabolism is discussed in Chapter 1.)
Diffusion Across the Alveolocapillary Membrane: The alveolocapillary membrane is the ideal medium for oxygen diffusion because it has a large total surface area (70 to 100 m2) and is very thin (0.5 μm). In addition, the partial pressure of oxygen molecules (PO2) is much greater in alveolar gas than in capillary blood, a condition that promotes rapid diffusion down the concentration gradient from the alveolus into the capillary.
The amount of oxygen in the alveoli (PAO2) depends on the amount of oxygen in the inspired air (see p. 1255) and on the amount of air that remains in the alveoli and tracheobronchial tree between breaths (physiologic dead space).1,4 This can be estimated by using the alveolar gas equation:

This value is approximately 104 with relaxed breathing; therefore a pressure gradient of approximately 60 mmHg facilitates the diffusion of oxygen from the alveolus into the capillary (Figure 32-17). Different values for PAO2 can be calculated if there are changes in the inspired oxygen content or the PaCO2, which are common occurrences in clinical settings.
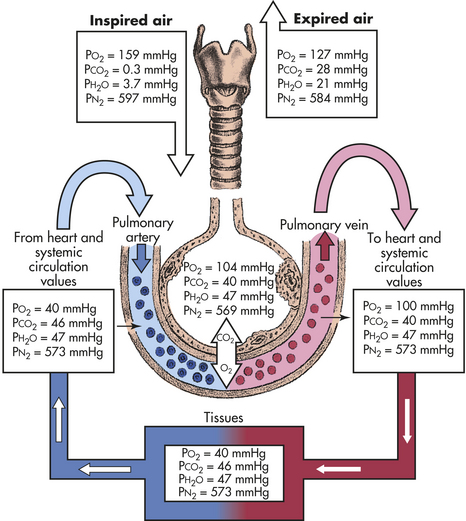
Figure 32-17 Partial pressure of respiratory gases in normal respiration. These are average values. The values of PO2, PCO2, and PN2 fluctuate from breath to breath. CO2, Carbon dioxide; H2O, water; N, nitrogen; O2, oxygen; PCO2, partial pressure of carbon dioxide; PH2O, partial pressure of water; PN2, partial pressure of nitrogen; PO2, partial pressure of oxygen.
Blood remains in the pulmonary capillary for about 0.75 second, but only 0.25 second is required for oxygen concentration to equilibrate (equalize) across the alveolocapillary membrane. Therefore, oxygen has ample time to diffuse into the blood, even during increased cardiac output, which speeds blood flow, shortening the time the blood remains in the capillary.
Determinants of Arterial Oxygenation: As oxygen diffuses across the alveolocapillary membrane, it dissolves in the plasma, where it exerts pressure (the partial pressure of oxygen in arterial blood, or PaO2). As the PaO2 increases, oxygen moves from the plasma into the red blood cells (erythrocytes) and binds with hemoglobin molecules. Oxygen continues to bind with hemoglobin until the hemoglobin binding sites are filled or saturated. Oxygen then continues to diffuse across the alveolocapillary membrane until the PaO2 and PAO2 equilibrate, eliminating the pressure gradient across the alveolocapillary membrane. At this point diffusion ceases (see Figure 32-17).
Normally approximately 20 ml of oxygen is transported per 100 ml of blood. Because oxygen is not very soluble in plasma, most of the oxygen molecules bind with hemoglobin. Plasma carries only about 0.3 ml of oxygen per 100 ml of blood (at sea level). Although the remaining 19.7 ml is carried by hemoglobin, it is the small amount of oxygen dissolved in plasma that is responsible for oxygen’s partial pressure (PaO2) in the blood.
Although PaO2 is important in that it provides the driving pressure that loads the hemoglobin with oxygen, it gives little information about the amount of oxygen carried in the blood. This amount, which is measured in milliliters per deciliter (100 ml) of blood, is the oxygen content of the blood. The total oxygen content of the blood depends on the amount of oxygen chemically combined with hemoglobin, as well as that dissolved in the blood. To calculate the total arterial oxygen content, we must know (1) hemoglobin concentration, or the amount of hemoglobin that is available to bind with oxygen (hemoglobin [Hb] in grams per deciliter); (2) the oxygen saturation or percentage of available hemoglobin that is bound to oxygen (SaO2); and (3) the partial pressure of oxygen (PaO2). The maximum amount of oxygen that can be transported by hemoglobin is 1.34 ml/g. The amount of oxygen that can be physically dissolved in blood is 0.003 ml/dl per mmHg. If these specific values are known, the oxygen content of arterial blood can be calculated.1
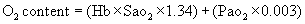
To calculate the oxygen content of venous blood, the partial pressure of mixed venous blood (PVO2) and venous oxygen saturation (SvO2) are substituted for the arterial values in the basic formula. Normal venous oxygen content is 15 to 16 ml/dl.
Because hemoglobin transports all but a small fraction of the oxygen carried in arterial blood, increases in hemoglobin concentration affect the oxygen content of the blood. Decreases in hemoglobin concentration below the normal value of 15 ml/dl of blood reduce oxygen content, and increases in hemoglobin concentration may minimize the effect of impaired gas exchange. In fact, an increase in hemoglobin concentration is a major compensatory mechanism in pulmonary diseases that impair gas exchange. For this reason, measurement of hemoglobin concentration is important in assessing individuals with pulmonary disease. If cardiovascular function is normal, the body’s initial response to low oxygen content is to speed up cardiac output. In individuals who also have cardiovascular disease, this compensatory mechanism does not work, making increased hemoglobin concentration an even more important compensatory mechanism. (Hemoglobin structure and function are described in Chapter 25.)
Oxyhemoglobin Association and Dissociation: When hemoglobin molecules bind with oxygen, oxyhemoglobin (HbO2) is formed. Binding occurs in the lungs and is called oxyhemoglobin association or hemoglobin saturation with oxygen (SaO2). The reverse process, in which oxygen is released from hemoglobin, occurs in the body tissues at the cellular level and is called hemoglobin desaturation. When hemoglobin saturation and desaturation are plotted on a graph, the result is a distinctive S-shaped curve known as the oxyhemoglobin dissociation curve (Figure 32-18).
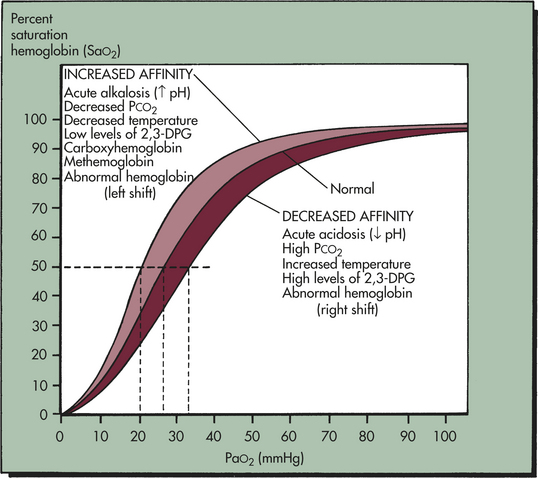
Figure 32-18 Oxyhemoglobin dissociation curve. The horizontal or flat segment of the curve at the top of the graph is sometimes called the arterial portion, or that part of the curve where oxygen is bound to hemoglobin. This portion of the curve is flat because partial pressure changes of oxygen between 60 and 100 mmHg do not significantly alter the percent saturation of hemoglobin with oxygen. The wide range of partial pressures of oxygen (PaO2—60 to 100 mmHg), represented by the flat part of the curve—allows adequate hemoglobin saturation at a variety of altitudes. For example, a PaO2 of 100 mmHg at sea level results in a hemoglobin saturation with oxygen of 98%. At an altitude of 5000 feet the PaO2 is about 70 mmHg and hemoglobin saturation is 94%, only 4% less than at sea level. If the relationship between SaO2 and PaO2 were linear (in a downward-sloping straight line) instead of flat between 60 and 100 mmHg, there would be inadequate saturation of hemoglobin with oxygen. For example, with a PaO2 of 70 mmHg the saturation would be only 70%, which is equivalent to normal venous oxygen saturation, and life could not be sustained at altitudes much above sea level. The steep part of the oxyhemoglobin dissociation curve occurs after the PaO2 drops below 60 mmHg and represents the rapid dissociation of oxygen from hemoglobin. During this phase oxygen diffuses rapidly from the blood into tissue cells. Conditions associated with altered affinity of hemoglobin for O2 are listed. P50 is the PaO2 at which hemoglobin is 50% saturated, normally 26.6 mmHg. A lower than normal P50 represents increased affinity of hemoglobin for O2; a high P50 is seen with decreased affinity. Note that variation from the normal is associated with decreased (low P50) or increased (high P50) availability of O2 to tissues (dotted lines). The shaded area shows the entire oxyhemoglobin dissociation curve under the same circumstances. 2,3-DPG, 2,3-diphosphoglycerate. (From Lane EE, Walker JF: Clinical arterial blood gas analysis, St Louis, 1987, Mosby.)
Several factors can change the relationship between PaO2 and SaO2, causing the oxyhemoglobin dissociation curve to shift to the right or left (see Figure 32-18). A shift to the right depicts hemoglobin’s decreased affinity for oxygen or an increase in the ease with which oxyhemoglobin dissociates and oxygen moves into the cells. A shift to the left depicts hemoglobin’s increased affinity for oxygen, which promotes association in the lungs and inhibits dissociation in the tissues.
The oxyhemoglobin dissociation curve is shifted to the right by acidosis (low pH) and hypercapnia (increased PaCO2). In the tissues the increased levels of CO2 and hydrogen ions produced by metabolic activity decrease the affinity of hemoglobin for oxygen. The curve is shifted to the left by alkalosis (high pH) and hypocapnia (decreased PaCO2). In the lungs, as CO2 diffuses from the blood into the alveoli, the blood CO2 level is reduced and the affinity of hemoglobin for oxygen is increased. The shift in the oxyhemoglobin dissociation curve caused by changes in CO2 and hydrogen ion concentration in the blood is called the Bohr effect.
The oxyhemoglobin curve is shifted also by changes in body temperature and increased or decreased levels of 2,3-diphosphoglycerate (2,3-DPG), a substance normally present in erythrocytes. Hyperthermia and increased 2,3-DPG levels shift the curve to the right. Hypothermia and decreased 2,3-DPG levels shift the curve to the left.
Carbon Dioxide Transport
Approximately 200 ml of CO2 is produced by the tissues per minute as a byproduct of cellular metabolism. This CO2 equilibrates with carbonic acid (H2O + CO2  H2CO3 H + HCO3−) and must be eliminated continuously to prevent acidosis. The elimination of CO2 by the lungs plays an important role in the regulation of acid-base balance (see Chapter 3).
H2CO3 H + HCO3−) and must be eliminated continuously to prevent acidosis. The elimination of CO2 by the lungs plays an important role in the regulation of acid-base balance (see Chapter 3).
CO2 is carried in the blood in three ways: (1) dissolved in plasma, (2) as bicarbonate, and (3) as carbamino compounds. As CO2 diffuses out of the cells into the blood, it dissolves in the plasma. Approximately 10% of the total CO2 in venous blood and 5% of the CO2 in arterial blood is carried dissolved in the plasma. As CO2 moves into the blood, it diffuses into the red blood cells. Within the red blood cells, CO2, with the help of the enzyme carbonic anhydrase, combines with water to form carbonic acid and then quickly dissociates into H+ and HCO3−. As carbonic acid dissociates, the H+ binds to hemoglobin, where it is buffered, and the HCO3− moves out of the red blood cell into the plasma. Approximately 60% of the CO2 in venous blood and 90% of the CO2 in arterial blood are carried in the form of bicarbonate. The remainder combines with blood proteins, hemoglobin in particular, to form carbamino compounds. Approximately 30% of the CO2 in venous blood and 5% of the CO2 in arterial blood are carried as carbamino compounds (see Figure 3-9).
CO2 is 20 times more soluble than O2 and diffuses quickly from the tissue cells into the blood. The amount of CO2 that is able to enter the blood is enhanced by diffusion of oxygen out of the blood and into the cells. Reduced hemoglobin (hemoglobin that is dissociated from oxygen) is able to carry more CO2 than hemoglobin that is saturated with O2. Therefore, the drop in SaO2 at the tissue level increases the ability of hemoglobin to carry CO2 back to the lung.
The diffusion gradient for CO2 in the lung is only approximately 6 mmHg (venous PvCO2 = 46 mmHg; alveolar PACO2 = 40 mmHg), yet CO2 is so soluble in the alveolocapillary membrane that the CO2 in the blood quickly diffuses into the alveoli, where it is removed from the lung with each expiration. Diffusion of CO2 in the lung is so efficient that diffusion defects that cause hypoxemia (low oxygen content of the blood) do not cause hypercapnia (excessive CO2 in the blood).
The diffusion of CO2 out of the blood also is enhanced by oxygen binding with hemoglobin in the lung. As hemoglobin binds with O2, the amount of CO2 carried by the blood is decreased. Thus in the tissue capillaries, O2 dissociation from hemoglobin facilitates the pickup of CO2, and the binding of O2 to hemoglobin in the lungs facilitates the release of CO2 from the blood. This effect of oxygen on CO2 transport is called the Haldane effect and can have significant clinical implications for the management of lung disease.10,11
Control of the Pulmonary Circulation
The caliber of pulmonary artery lumina decreases as smooth muscle in arterial walls contracts. Contraction increases pulmonary artery pressure. Caliber increases as these muscles relax, decreasing blood pressure. Contraction (vasoconstriction) and relaxation (vasodilation) apparently occur in response to local humoral conditions, even though the pulmonary circulation is innervated by the ANS in the same manner as the systemic circulation.
The most important cause of pulmonary artery constriction is a low alveolar partial pressure of oxygen (PAO2). Vasoconstriction caused by alveolar and pulmonary venous hypoxia, often termed hypoxic pulmonary vasoconstriction, can affect only one portion of the lung or the entire lung. If only one segment of the lung is involved, the arterioles to that segment constrict, shunting blood to other, well-ventilated portions of the lung. This reflex improves the lung’s efficiency by better matching ventilation and perfusion. If alveolar hypoxia affects all segments of the lung, however, vasoconstriction occurs throughout the pulmonary vasculature, and pulmonary hypertension (elevated pulmonary artery pressure) can result. The pulmonary vasoconstriction caused by low PAO2 is reversible if the PACO2 is corrected. Chronic alveolar hypoxia
can result in permanent pulmonary artery hypertension, which eventually leads to cor pulmonale and heart failure (see What’s New? Hypoxic Pulmonary Vasoconstriction).12,13
Acidemia also causes pulmonary artery constriction. If the acidemia is corrected, the vasoconstriction is reversed. (Respiratory acidosis and metabolic acidosis are described in Chapter 3.) It is important to note that an elevated PaCO2 without a drop in pH does not cause pulmonary artery constriction. Other biochemical factors that affect the caliber of vessels in pulmonary circulation are histamine, prostaglandins, endothelin, serotonin, nitric oxide, and bradykinin.
TESTS OF PULMONARY FUNCTION
Several laboratory tests aid in the diagnosis and evaluation of pulmonary system abnormalities. Most of them are easy to perform at hospitals and clinics. They provide valuable information as to the possible cause of a respiratory abnormality and evaluate the progression or resolution of disease.
Spirometry is used to measure forced expiration, which often is affected by diffuse pulmonary disease. Because the pulmonary system has remarkable reserves, disease may become well established before clinical manifestations appear. Spirometry enables clinicians to detect restrictive or obstructive deficits early in the course of disease. Restrictive lung diseases restrict the lungs’ volume: the lungs are unable to expand normally, diminishing the amount of gas that can be inspired. Obstructive diseases affect gas flow: airflow into and out of the lungs is obstructed.
Spirometry measures both volume and flow. The test is performed with a spirometer, which is a water-filled cylinder into which an inverted cylinder or bell has been inserted. A length of tubing runs from the inverted bell to a mouthpiece through which a person breathes during testing. The bell is attached to a pen that writes on calibrated paper rotating at a constant speed. As a person performs various breathing maneuvers, the inverted bell moves up and down, causing the pen to move on the calibrated paper. This produces a spirogram, which is a record of the individual’s ventilation in relation to time (Figure 32-19). Clinically the most important spirometric tests are the forced vital capacity (FVC) and the forced expiratory volume in one second (FEV1).13 (These tests and other important measures are described in Table 32-3.)
Table 32-3
| Symbol | Ventilatory Property Measured |
| FVC | Forced vital capacity: maximum amount of gas that can be displaced from the lung during a forced expiration |
| FEV1 | Forced expiratory volume in 1 second: maximum amount of air that can be expired from the lung in 1 second |
| FEV1/FVC | Percentage of maximum inspiration that is expired in 1 second, usually 80% of FVC |
| FEV3 | Forced expiratory volume in 3 seconds; maximum amount of gas that can be expired in 3 seconds |
| FEV3/FVC | Percentage of FVC that is expired in 3 seconds; usually 95% of FVC |
| FEF25%-75% | Forced expiratory flow rate during the middle 50% of expiration; sometimes reported as maximum midexpiratory flow rate (MMFR) |
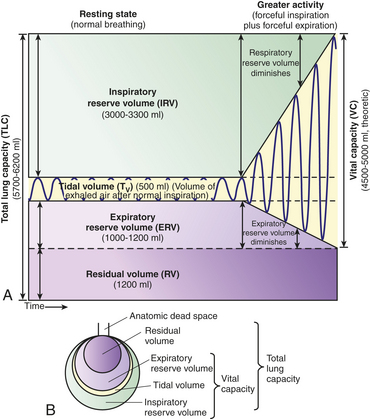
Figure 32-19 Pulmonary ventilation and lung capacities. A, Spirogram. During normal, quiet respirations the atmosphere and lungs exchange about 500 ml of air (VT). With a forcible inspiration, about 3300 ml more air can be inhaled (IRV). After a normal inspiration and normal expiration, approximately 1000 ml more air can be forcibly expired (ERV). Vital capacity is the amount of air that can be forcibly expired after a maximal inspiration and indicates, therefore, the largest amount of air that can enter and leave the lungs during respiration. Residual volume is the air that remains trapped in the alveoli. B, Lung capacities. (From, Patton KT, Thibodeau GA Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
Lung capacities, such as vital capacity and total lung capacity, are always the sum of two or more volumes. Norms for volumes and capacities are based on age, gender, and height and are referred to as predicted values. Changes from predicted or baseline values are taken into account in diagnosing and assessing respiratory disorders.
Diffusing capacity is a measure of the rate of gas diffusion across the alveolocapillary membrane. Oxygen, or more commonly carbon monoxide, is used to measure diffusing capacity. The measurement is made by determining how much carbon monoxide is taken up by the blood and dividing this amount by the pressure gradient across the alveolocapillary membrane. Helium often is added to the gas mixture to obtain a simultaneous measurement of residual volume (RV), functional reserve capacity (FRC), and total lung capacity (TLC). Individuals are asked to perform ventilatory maneuvers similar to those of spirometry. A decreased diffusing capacity can be the result of an abnormal ventilation-perfusion ratio or an actual diffusion defect. Diffusing capacity is decreased in individuals with emphysema.
Arterial blood gas analysis commonly is performed for individuals with suggested or diagnosed pulmonary disease. Direct analysis of the pH and gas concentrations in arterial blood provides valuable information about an individual’s gas exchange and acid-base status. Acidosis (low pH), alkalosis (high pH), ventilatory alterations, and decreased PaO2 can be diagnosed accurately only by arterial blood gas analysis. A blood gas report may be divided into an acid-base/ventilation portion and an oxygenation portion. (Normal values for arterial blood gases are given in Table 32-4; acid-base alterations are described in Chapter 3.) Oximetry can be used to monitor oxygen saturation once the arterial blood gas analysis has accurately measured the PaO2, but it does not measure PaCO2 or pH.
Table 32-4
Normal Ranges for Arterial and Mixed Venous Blood Gases
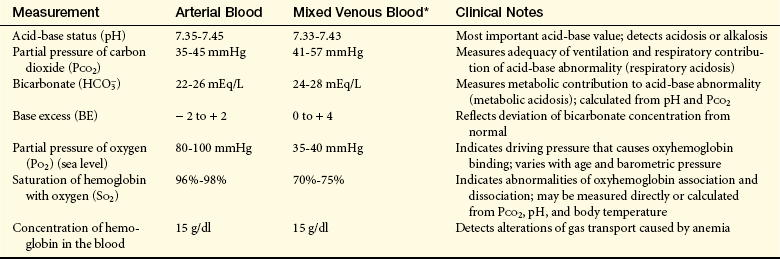
∗Mixed venous (pulmonary artery) blood is analyzed for critically ill individuals and those undergoing cardiac catheterization (it is not practical to withdraw samples except from a pulmonary artery catheter). Mixed venous blood gas analysis, in conjunction with arterial analysis, provides important information about the adequacy of cardiac output and tissue oxygenation.
Signs and symptoms of most respiratory abnormalities first appear when the system is stressed during exercise. Therefore, if pulmonary disease is suspected, the individual is evaluated at rest and during exercise. During exercise the usual procedures are spirometry and withdrawal of arterial blood for gas analysis. The exercise usually consists of riding a stationary bicycle or walking on a treadmill. Exercise testing enables clinicians to detect early changes in respiratory function and thus begin treatment. Exercise tests also are used in planning and evaluating exercise and rehabilitation programs.
Chest radiographs are among the most common examinations of the pulmonary system. A few of the abnormalities detected in chest radiographs are air trapping in the alveoli and airways (e.g., in asthma or emphysema), consolidation of lung tissue (in pneumonia or pulmonary edema), cavities (abscesses or tuberculosis), and nodules (lung cancer). Often pulmonary abnormalities are detected in routine chest radiographs of asymptomatic individuals. Various radiographic techniques are available for the diagnosis and evaluation of respiratory disorders.
Aging and the Pulmonary System
Most knowledge about pulmonary structure and function is based on norms for the middle years. Less is known about structure and function in the very young (see Chapter 34) and older adults, but a few normal physiologic (developmental and degenerative) changes are known to occur from birth to old age. An understanding of these changes is needed to provide appropriate care and to differentiate between normal alterations and disease. Normal alterations include (1) loss of elastic recoil, (2) stiffening of the chest wall, (3) alterations in gas exchange, and (4) increases in flow resistance (Figure 32-20). These changes are influenced by environmental factors, respiratory disease, body size, and race.14
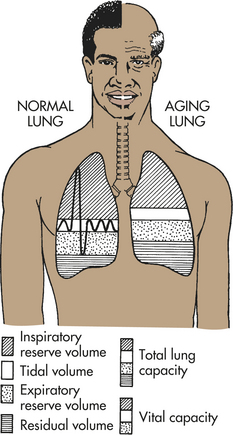
Figure 32-20 Changes in lung volumes with aging. With aging, note particularly the decrease in vital capacity and the increase in residual volume.
During adulthood and as age advances, the alveoli tend to lose alveoli wall tissue and capillaries. This process diminishes alveolar surface area available for gas diffusion and decreases airway support provided by normal lung tissues. Mechanical changes involve elastic properties of the lungs and chest wall. Chest wall compliance decreases with age because the ribs become ossified (less flexible) and joints become stiffer. As a result the chest wall loses some of its ability to expand. In addition, respiratory muscle strength and endurance decrease by up to 20% by age 70.14 These mechanical changes in the lung and chest wall, along with structural changes in the alveoli, reduce ventilatory capacity in older adults. Vital capacity decreases and residual volume increases; however, total lung capacity remains unchanged. These changes decrease ventilatory reserves and lead to decreased ventilation-perfusion ratios.14 With advancing age there is also increased immune dysregulation, asymptomatic low-grade inflammation, and increased risk of infection.14–16
Alterations in gas exchange are reflected by blood gas analysis. With advancing age, pH and PaCO2 do not change much, even though it has been documented that the chemoreceptors become less sensitive to gas partial pressures with age. Older adults have a decreased compensatory response to hypercapnia and hypoxemia; however, the perception of dyspnea remains intact and is even enhanced. PaO2 declines with age as a result of structural and mechanical changes, such as loss of alveolar surface area and increased ventilation-perfusion mismatch. The maximum PaO2 in an older adult at sea level can be estimated by multiplying the person’s age by 0.3 and subtracting the product from 100. For example, an 80-year-old individual would have an estimated maximum PaO2 of 76 mmHg (0.3 × 80 = 24; 100 − 24 = 76).17 There is also a decrease in the capillary network.
The decrease in PaO2 and diminished ventilatory reserve in an older adult lead to a decrease in exercise tolerance. Respiratory muscle strength and endurance decrease with age.17 Furthermore, older adults are at greater risk for respiratory depression caused by medications. Changes in respiratory function can vary considerably from person to person, however. Changes also are affected by activity and fitness earlier in life. A very active, physically fit individual will, all else being equal, have fewer changes in function at any age than one who has been sedentary.
Airway resistance 1253
Acinus 1244
Alveolar duct 1244
Alveolar ventilation 1249
Alveoli (sing., alveolus) 1244
Alveolocapillary membrane 1247
Arterial blood gas analysis 1262
Bohr effect 1259
Bronchiole 1244
Bronchoconstriction 1254
Bronchodilation 1254
Bronchus (pl., bronchi) 1244
Carina 1244
Central chemoreceptor 1251
Chest radiograph 1262
Compliance 1253
Diaphragm 1252
Diffusing capacity 1262
Elastic recoil 1253
Functional reserve capacity (FRC) 1262
Gas transport 1255
Goblet cell 1244
Haldane effect 1260
Hilus (pl., hila) 1244
Hypoxic pulmonary vasoconstriction 1260
Irritant receptor 1251
J-receptor 1251
Larynx 1243
Lymphatic capillary 1249
Minute volume 1249
Nasopharynx 1242
Oropharynx 1242
Oxygen content 1258
Oxyhemoglobin (HbO2) 1259
Oxyhemoglobin dissociation curve 1259
Partial pressure (of a gas) 1255
Peripheral chemoreceptor 1250, 1251
Physiologic dead space 1257
Pleura (pl., pleurae) 1249
Pleural space (pleural cavity) 1249
Pulmonary artery 1247
Pulmonary capillary 1247
Pulmonary vein 1247
Rapidly adapting receptor (RAR) 1251
Residual volume (RV) 1262
Respiration 1249
Respiratory bronchiole 1244
Respiratory center 1250
Spirometry 1261
Stretch receptor 1251
Surface tension 1252
Surfactant 1244
Thoracic cavity 1249
Total lung capacity (TLC) 1262
Trachea 1244
Ventilation 1249
Ventilation-perfusion ratio () 1257
Work of breathing 1254
REFERENCES
1. Lumb, A.B. Nunn’s applied respiratory physiology, ed 6. St Louis: Elsevier; 2005.
2. Ganong, W. Review of medical physiology, ed 22. Philadelphia: McGraw-Hill; 2005.
3. Corrin, B. Pathology of the lungs. London: Churchill Livingstone; 2000.
4. West, J.B. Respiratory physiology: the essentials, ed 8. Philadelphia: Lippincott Williams & Wilkins; 2008.
5. Benarroch, E.E. Brainstem respiratory chemosensitivity: new insights and clinical implications. Neurology. 2007;68(24):2140–2143.
6. Haberthur, C., Guttmann, J. Short-term effects of positive end-expiratory pressure on breathing pattern: an interventional study in adult intensive care patients. Crit Care. 2005;9(4):R407–R415.
7. Canning, B.J. Anatomy and neurophysiology of the cough reflex. Chest. 2006;129:33s–47s.
8. Kuroki, Y., Takahashi, M., Nishitani, C. Pulmonary collectins in innate immunity of the lung. Cell Microbiol. 2007;9(8):1871–1879.
9. Des Jardins, T., Burton, G.C. Clinical manifestations and assessment of respiratory disease, ed 5. St Louis: Mosby; 2006.
10. Cavaliere, F., et al. Comparison of two methods to assess blood CO2 equilibration curve in mechanically ventilated patients. Resp Physiol Neurobiol. 2005;146(1):77–83.
11. Chien, J.W., et al. Uncontrolled oxygen administration and respiratory failure in acute asthma. Chest. 2000;117(3):728–733.
12. Pak, O., et al. The effects of hypoxia on the cells of the pulmonary vasculature. Eur Resp J. 2007;30(2):364–372.
13. Mason R.J., et al, eds. Murray and Nadel’s textbook of respiratory medicine, ed 4, St Louis: Elsevier, 2005.
14. Sharma, G., Goodwin, J. Effect of aging on respiratory system physiology and immunology. Clin Interven Aging. 2006;1(3):253–260.
15. Haas, C.F., Loik, P.S., Gay, S.E. Airway clearance applications in the elderly and in patients with neurologic or neuromuscular compromise. Respir Care. 2007;52(10):1362–1381. [discussion 1381].
16. Ely, K.H., et al. Aging and CD8+ T cell immunity to respiratory virus infections. Exp Gerontol. 2007;42(5):427–431.
17. Hardie, J.A., et al. Reference values for arterial blood gases in the elderly. Chest. 2004;125(6):2053–2060.