STRUCTURE AND FUNCTION OF THE MUSCULOSKELETAL SYSTEM


The way an individual functions in daily life, moves about, or manipulates objects physically depends on the integrity of the musculoskeletal system. The musculoskeletal system is actually composed of two systems: (1) the skeleton proper, which is composed of bones and joints; and (2) skeletal muscles. Each of the systems contributes to mobility. The skeleton supports the body and provides leverage to the skeletal muscles so that movement of various parts of the body is possible. This movement is accomplished by contraction of the skeletal muscles and bending or rotation at the joints.
STRUCTURE AND FUNCTION OF BONES
Bones give form to the body, support tissues, and permit movement by providing points of attachment for muscles. Many bones meet in movable joints that determine the type and extent of movement possible. Bones also protect many of the body’s vital organs. For example, the bones of the skull, thorax, and pelvis are hard exterior shields that protect the brain, heart, lungs, and reproductive and urinary organs.
The marrow cavities within certain bones serve as sites of blood cell formation. In adults, blood cells originate exclusively in the marrow cavities of the skull, vertebrae, ribs, sternum, shoulders, and pelvis. Bones also have a crucial role in mineral homeostasis, and storing and releasing minerals (i.e., calcium, phosphate, carbonate, magnesium) that are essential for the proper working of many delicate cellular mechanisms.
Elements of Bone Tissue
Mature bone is a rigid yet flexible connective tissue consisting of cells, fibers, a gelatinous material termed ground substance, and large amounts of crystallized minerals, mainly calcium, that give bone its rigidity. The structural elements of bone are summarized in Table 41-1.
Table 41-1
| Structural Element | Function |
| Bone Cells | |
| Osteoblasts | Synthesize collagen and proteoglycans; stimulate bone formation and are also involved in some osteoclast resorptive activity |
| Osteocytes | Maintain bone matrix; act as mechanoreceptors, influence osteoblasts and osteoclasts |
| Osteoclasts | Resorb bone; assist with mineral homeostasis |
| Bone Matrix | |
| Collagen fibers | Lend support and tensile strength |
| Proteoglycans | Control transport of ionized materials through matrix |
| Bone morphogenic proteins (BMPs)∗ | Induce cartilage and bone formation |
| BMP-2 | Induces osteoblast differentiation in mesenchymal stem cells |
| BMP-6 | Accelerates bone repair |
| BMP-9 | Induces osteogenesis in mature osteoblasts |
| Glycoproteins | |
| Sialoprotein | Promotes calcification |
| Osteocalcin | Inhibits calcium-phosphate precipitation; promotes bone resorption |
| Laminin | Stabilizes basement membranes in bones |
| Osteonectin | Binds calcium in bones |
| Albumin | Transports essential elements to matrix; maintains osmotic pressure of bone fluid |
| α-Glycoprotein | Promotes calcification |
| Minerals (elements) | Crystallizes to lend rigidity and compressive strength |
| Calcium | Regulates vitamin D and thereby promotes mineralization |
| Phosphate |
∗Data from Caetano-Lopes J, Canhão H, Fonseca JE: Arthritis Res Ther 9(Suppl 1):S1, 2007.
Bone cells enable bone to grow, repair itself, change shape, and continuously synthesize new bone tissue and resorb (dissolve or digest) old tissue. The fibers in bone are made of collagen, which gives bone its tensile strength (the ability to hold itself together). Ground substance acts as a medium for the diffusion of nutrients, oxygen, metabolic wastes, biochemicals, and minerals between bone tissue and blood vessels.
Bone formation begins during embryonic development when mesenchymal stem cells begin differentiating into either chondrocytes or preosteoblasts. Endochondral ossification and intramembranous bone formation are the two major mechanisms responsible for normal bone development.
Endochondral ossification occurs when mesenchymal (mesenchyme, or loose tissue found during embryonic development) stem cells begin differentiating into chondrocytes (Figure 41-1), which in turn develop a mineralized cartilage scaffold that allows formation of osteoblasts. Most bone elements are formed this way. With the second mechanism, intramembranous bone formation, mesenchymal stem cells differentiate into a preosteoblast line that then forms osteoblasts without any cartilage framework.1,2
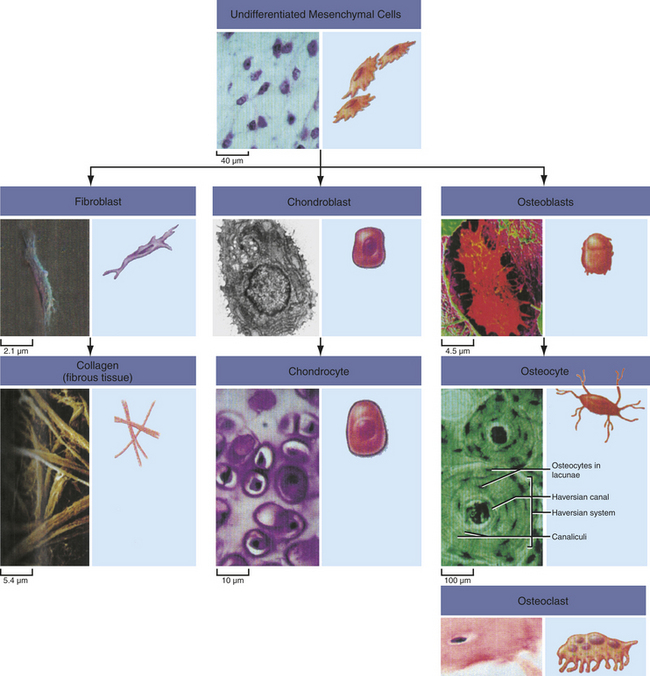
Figure 41-1 Mesenchymal stem cells. Undifferentiated mesenchymal cells give rise to a variety of cell types with distinct functions. (From Raven PH et al, editors: Biology, ed 8, New York, 2008, McGraw-Hill.)
One of the major genetic factors influencing normal bone development and growth is a signaling family called Wnts. Wnt signaling of chondrocytes and osteoblasts affects bone mass and density, joint formation, fracture repair, and bone remodeling, as well as some bone diseases.3 Osteoblasts are the principal cellular targets of Wnt signaling in bone.4
Wnt genes belong to a large family of protein-signaling factors that are required for the development of body systems, including the musculoskeletal system. They play a significant role in bone formation, developing bone mass, remodeling, and fracture healing. Wnt signaling regulates production and differentiation of osteoblasts and osteoclasts.1,3,5
Another major influence on bone and cartilage development is bone morphogenic proteins (BMPs). BMPs are multifunctional growth factors that belong to the transforming growth factor-beta (TGF-β) superfamily and affect several systems, including embryonic and postnatal cartilage and bone development. BMP activities are regulated at different molecular levels. Table 41-2 summarizes the role of several important BMP proteins.
Table 41-2
Bone Morphogenic Proteins (BMPs) and Their Functions
| BMP | Known Function |
| BMP1 | Cartilage development; is actually a metalloprotease |
| BMP2 | Induces bone and cartilage formation |
| BMP3 | Induces bone formation |
| BMP4 | Regulates formation of teeth, limbs, and bone |
| BMP5 | Involved in cartilage development |
| BMP6 | Helps maintain adult joint integrity |
| BMP7 | Major role in osteoblast differentiation; important in renal development and repair |
| BMP8a | Bone and cartilage development |
| BMP8b | Found in hippocampus |
| BMP10 | May play role in development of the heart |
In mature bone the formation of new tissue begins with the production of an organic matrix by the bone cells. This bone matrix consists of ground substances, collagen, and other proteins (see Table 41-1) that take part in bone formation and maintenance.
The next step in bone formation is calcification, when minerals are deposited and crystallize. Minerals bind tightly to collagen fibers, producing tensile and compressional strength in bone, and withstand pressure and weightbearing.
Bone Cells
Bone contains three types of cells: osteoblasts, osteocytes, and osteoclasts (Figure 41-2). Osteoblasts are the bone-forming cells, whose primary function is to lay down new bone. Once this function is complete, osteoblasts become osteocytes. Osteocytes are the most plentiful cells in bone. Osteocytes are osteoblasts that have become imprisoned within the mineralized bone matrix. They help maintain bone by signaling osteoblasts and osteoclasts to form and resorb bone.6
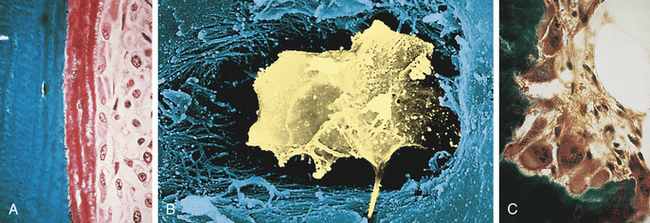
Figure 41-2 Bone cells. A, Osteoblasts are responsible for the production of collagenous and noncollagenous proteins that compose osteoid. Active osteoblasts are lined up on the osteoid. Note the eccentrically located nuclei. B, Osteocyte. Scanning electron micrograph showing an osteocyte within a lacuna. The cell is surrounded by collagen fibers and mineralized bone. C, Osteoclasts actively resorb mineralized tissue. The scalloped surface in which the multinucleated osteoclasts rest is termed the Howship lacuna. (A and C from Damjanov I, Linder J, editors: Anderson’s pathology, ed 10, St Louis, 1996, Mosby. B from Erlandsen S, Magney J: Color atlas of histology, St Louis, 1992, Mosby.)
Osteoblasts: Osteoblasts are cells derived from mesenchymal stem cells. Osteoblasts produce several substances, including osteocalcin, TGF-β (a growth inhibitor for many cells), macrophage colony-stimulating factor, receptor activator of nuclear factor κB-ligand, osteoprotegerin (OPG), and bone matrix7 and osteocalcin when stimulated by 1,25-dihydroxyvitamin D. Osteoblasts are active on the outer surface of bones, where they form a single layer of cells. They bring about the formation of new bone by their synthesis of osteoid (nonmineralized bone matrix). The mechanism of osteoblast stimulation is reported to be the production of so-called coupling factors generated during the resorption process. Osteoblasts bring about the formation of new bone and the orderly mineralization of bone matrix by concentrating some of the plasma proteins (growth factors) found in the bone matrix and by facilitating the deposit and exchange of calcium and other ions at the site. Osteoblasts alter levels of receptor activator of nuclear factor κB-ligand (RANKL) and OPG. The balance of these two cytokines determines overall osteoclast formation8 (see Figure 42-11). RANKL is part of a ligand-receptor system that assists in regulating osteoclastic bone remodeling. As new bone is formed it is shaped and remodeled through TGF-β, as well as other plasma proteins (growth factors) found in the bone marrow (Table 41-3).
Table 41-3
Effects of Selected Cytokines (Growth Factors) on Skeletal Tissues
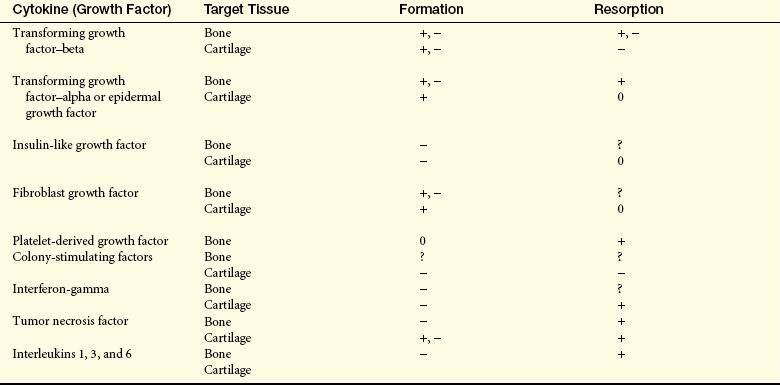
+, −, Both stimulatory and inhibitory properties on the specific cell listed; 0, no effects presently known; ?, possible effects on cell listed.
Osteocytes communicate with other cells and appear to instruct both osteoclasts and osteoblasts about when and where to resorb and form bone.6 In contact with bone mineral, osteoclasts use adhesive structures, called podosomes, to attach themselves to the bone surface. They then seal their cytoplasm with the underlying bone and dissolve the bone and matrix while protecting the surrounding tissue.9 Thus the cells of the osteoblastic lineage (osteoblasts and osteocytes) form a network of cells in bone that sense the shape and structure of bone and determine where it is appropriate that bone be formed or resorbed, according to Wolfe’s law (bone is shaped according to its function).
Osteoblasts have an active state and a resting state. When active, osteoblasts synthesize and secrete osteoid. When in the resting state, they appear dormant. If appropriately stimulated, however, the resting osteoblasts are capable of resuming activity.
Osteocytes: An osteocyte is a transformed osteoblast that is trapped or surrounded in osteoid as it hardens from minerals that enter during ossification (see Figure 41-2, B). It is the final differentiation stage for an osteoblast. The osteocyte is within a space in the hardened bone matrix called a lacuna. Each osteocyte has a high nucleus/cytoplasm ratio with a thin layer of nonmineralized osteoid around it, similar to the egg white surrounding an egg yolk. Osteocytes secrete proteins, such as sclerostin (a Wnt antagonist that binds BMPs), that result in reduced bone formation.4,7
The function of osteocytes is not fully known, but they communicate with each other and help concentrate nutrients in the matrix. Osteocytes obtain nutrients from capillaries in the canaliculi, which contain nutrient-rich fluids. Osteocytes also help synthesize and replace needed elements of the matrix by signaling both osteoclasts and osteoblasts to resorb and form new bone. Through exchanges between these cells, hormone catalysts, and minerals, optimal levels of calcium, phosphorus, and other minerals are maintained in blood plasma. The osteocyte also aids in modifying bone matrix through release of enzymes to dissolve the mineralized walls of the lacunae to prepare the bone for remodeling. (Remodeling is described on p. 1547.)
Osteoclasts: Osteoclasts are the major resorptive cells of bone. They are large multinucleated cells with a short life span. Osteoclasts develop from the hematopoietic stem cell in the bone marrow stroma8 and adjacent vessels and from mononuclear phagocytic cells. Osteoclasts contain lysosomes (digestive vacuoles) filled with hydrolytic enzymes. Fine projections, or microvilli, fan out from the osteoclast cell’s surface and are known as ruffled borders; these projections result from extensive infoldings of the cell membrane adjacent to the resorptive surface.10 Osteoclasts in regions of bone resorption lie in pits called Howship lacunae, where the infolded, ruffled borders of the osteoclasts greatly increase the surface area of the plasma membrane. The infolds end in numerous channels and vesicles in the cell cytoplasm, permitting them to resorb the bone under their ruffled, infolded borders.
Osteoclasts bind to the bone surface of cell attachment proteins called integrins.10 They bring about resorption of bone by secretion of hydrochloric acid (HCl) and cathepsin K (a protease enzyme),9 which help dissolve bone minerals and collagenase, which aids in digesting collagen, along with the action of cytokines (see Table 41-3). Matrix metalloproteinases (MMPs), a group of proteolytic enzymes, help control osteoclast-matrix interactions necessary for bone resorption.11 Once resorption is completed, the osteoclast disappears by degeneration, either by reverting back to its parent cell or by leaving the site through the process of cell mobility, wherein the osteoclast then becomes an inactive, or “resting,” osteoclast.
Bone Matrix
Bone matrix is made of the extracellular elements of bone tissue, composed of about 35% organic and 65% inorganic materials. The major organic components are collagen fibers, and the major inorganic components are the calcium and phosphate minerals. Other parts of the bone matrix are the proteins, carbohydrate-protein complexes, and ground substances. Water makes up 5% to 8% of the matrix.
Collagen Fibers: Collagen fibers are the major organic component of bone matrix. The fibers are approximately 90% type I collagen, which is synthesized and secreted by osteoblasts. Collagen-I is essential for bone strength. Once secreted, the collagen molecules assemble into thin chains called α-chains, which combine in threes to form fibrils. The fibrils form a staggered pattern, overlapping nearby fibrils by approximately one fourth their length. This staggered, overlapping pattern creates regular gaps, called hole zones, into which mineral crystals are deposited. After mineral deposition the fibrils link together and twist to form ropelike fibers. Collagen fibers then join to form a framework that gives bone its tensile strength and enables it to bear weight.
Collagen is the most abundant macromolecule in the body, accounting for approximately one third of all protein and providing the structural framework for nearly all tissues. Collagen is one of the extracellular components, along with proteoglycans and noncollagenous matrix proteins, of articular cartilage. To date, several types of collagen have been identified. Cartilage-specific collagens include types II (the principal component), VI, IX, X, and XI. Type IX collagen is thought to be the “glue” that holds together the type II collagen scaffold of articular cartilage, helps maintain the structural integrity of cartilage, and resists tensile forces on the joint cartilage. Type XI regulates the fibril diameter of type II cartilage. Degradation of type IX collagen by proteolytic enzymes has been seen in the early stages of osteoarthritis and rheumatoid arthritis. Researchers have proposed that this degradation, or “unplugging,” may be the mechanism for the degenerative changes seen in osteoarthritic and rheumatoid cartilage. Table 41-4 gives the musculoskeletal distribution of other types of collagen.
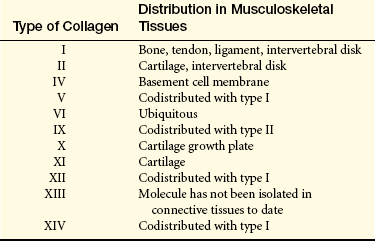
Proteoglycans: Proteoglycans are large complexes of numerous polysaccharides attached to a common protein core. Hyaline cartilage is primarily composed of the glycosaminoglycans chondroitin sulfate and keratin sulfate. They strengthen bone by forming compression-resistant networks between the collagen fibrils. Proteoglycans also control the transport and distribution of electrically charged particles (ions), particularly calcium, through the bone matrix, thereby playing a role in bone calcium deposition and calcification.
Glycoproteins: Glycoproteins are also carbohydrate-protein complexes of bone. Glycoproteins control the collagen interactions that lead to fibril formation. They also may play a role in calcification.
Some of the glycoproteins in bone matrix include bone sialoprotein, osteocalcin, osteonectin, laminin, albumin, and α-glycoprotein. Other proteins found in bone matrix are the compounds currently called bone morphogenic proteins (BMPs) (see Table 41-1). Sialoprotein (osteopontin) makes up about 8% of the noncollagenous matrix of bone and easily binds with calcium.
Osteocalcin is also a calcium-binding protein that binds preferentially to calcium that has already crystallized. The roles of osteocalcin may be to inhibit calcium phosphate precipitation and play a part in bone resorption by recruiting osteoclasts.
Osteonectin is also thought to bind calcium in bones, and laminin stabilizes basement membranes in bones. Laminin is an abundant bone matrix protein in humans that is most effective in neurite and axon growth.
Bone albumin is identical to serum albumin. In calcified matrix, bone albumin is permanently fixed to bone mineral crystals and remains so until the bone is resorbed. Researchers believe bone albumin transports essential elements such as hormones, ions, and other metabolites to and from the bone cells and maintains the osmotic pressure of bone fluid (fluid surrounding mineral crystals and osteoblasts).
α-Glycoprotein is thought to be synthesized in the liver, to be released into blood plasma, and to circulate to bone matrix, where it accumulates. α-Glycoprotein’s affinity for calcium is 40 times greater than that of albumin. Therefore, it probably plays an important role in the calcification of growing bone. α-Glycoprotein also may facilitate bone resorption by activating osteoclasts.
Bone Minerals
Mineralization is the final step in bone formation, after collagen synthesis and fiber formation. Mineralization has two distinct phases: (1) formation of the initial mineral deposit (initiation), and (2) proliferation or accretion of additional mineral crystals on the initial mineral deposits (growth).10 The majority of the mineral content in the body is an analog of the naturally occurring mineral hydroxyapatite.
Table 41-5 lists the sequence in which calcium and phosphate form amorphous (fluid) calcium phosphate compounds that are converted, in stages, to solid hexagonal crystals of hydroxyapatite (HAP). As the calcium and phosphorus concentrations increase in the bone matrix, the first precipitate to form is dicalcium phosphate dihydrate (DCPD). Once DCPD precipitation begins, the remaining phases of bone crystal formation proceed until insoluble HAP is produced, with approximately 80% to 90% of the HAP being incorporated into the collagen fibers. Amorphous calcium phosphate is distributed throughout the bone matrix.
Table 41-5
Sequence of Calcium and Phosphate Compound Formation and Crystallization
| Formula | Name | Abbreviation |
| Ca(HPO4) × 2H2O | Dicalcium phosphate dihydrate | DCPD |
| Ca4H(PO4)3 | Octacalcium phosphate | OCP |
| Ca9(PO4)6 (var.) | Amorphous calcium phosphate | ACP |
| Ca3(PO4)2 | Tricalcium phosphate | TCP |
| Ca5(PO4)3OH | Hydroxyapatite | HAP |
NOTE: Compounds are listed in the order in which precipitation and crystal formation occur.
Types of Bone Tissue
Bone is made up of two types of bony (osseous) tissue: compact bone (cortical bone) and spongy bone (cancellous bone) (Figure 41-3). Compact bone makes up approximately 85% of the skeleton; spongy bone makes up the remaining 15%. Both types of bone tissue contain the same structural elements, and, with a few exceptions, both compact tissue and spongy tissue are present in every bone. The major difference between the two types of tissue is the organization of the elements.
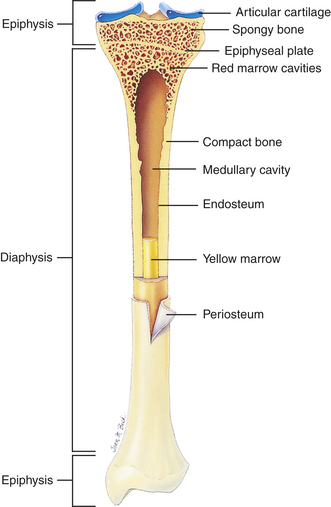
Figure 41-3 Cross section of bone. Longitudinal section of long bone (tibia) showing cancellous and compact bone. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
Compact bone is highly organized, solid, and extremely strong. The basic structural unit in compact bone is the haversian system (Figure 41-4). Each haversian system is made up of the following:
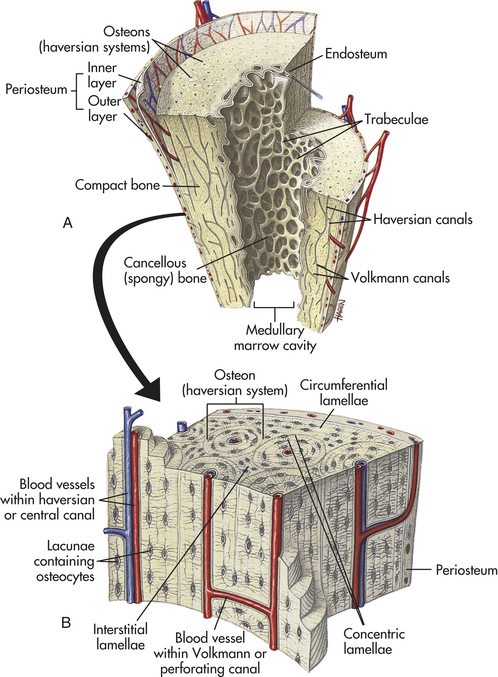
Figure 41-4 Structure of compact and cancellous bone. A, Longitudinal section of a long bone showing cancellous and compact bone. B, A magnified view of compact bone. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
1. A central canal called the haversian canal
2. Concentric layers of bone matrix called lamellae
3. Tiny spaces (lacunae) between the lamellae
Each haversian system is a separate cylindrical entity that looks like a set of concentric rings. In the center of the haversian system is the haversian canal. The haversian canal runs through the long axis of bone and contains one or two blood vessels and nerve fibers. The blood vessels in the canal communicate with blood vessels in the periosteum (surface cover) and marrow cavity to transport nutrients and wastes to and from the osteocytes contained within the lacunae. Surrounding each haversian canal are the concentric lamellae. Between the lamellae are the lacunae, each of which contains one osteocyte. The lacunae are connected to each other and to the haversian canal by the canaliculi, which run parallel to the horizontal axis of the bone. Each canaliculus encloses a small extension (cytoplasmic process) from the osteocyte contained in the lacuna. The canaliculi transport both nourishment and molecular signals into the lacunae, a mechanism essential for osteocyte survival.
Spongy bone is less complex and lacks haversian systems. In spongy bone the lamellae are not arranged in concentric layers but in plates or bars termed trabeculae that branch and unite with one another to form an irregular meshwork. The pattern of the meshwork is determined by the direction of stress on the particular bone. The spaces between the trabeculae are filled with red bone marrow. The osteocyte-containing lacunae are distributed between the trabeculae and interconnected by canaliculi. Capillaries pass through the marrow to nourish the osteocytes.
All bones are covered with a double-layered connective tissue called the periosteum. The outer layer of the periosteum contains blood vessels and nerves, some of which penetrate to the inner structures of the bone through channels called Volkmann canals (see Figure 41-4). The inner layer of the periosteum is anchored to the bone by collagenous fibers (Sharpey fibers) that penetrate the bone. Sharpey fibers also help hold or attach tendons and ligaments to the periosteum of bones.10
Characteristics of Bone
The human skeleton consists of 206 bones that constitute the axial skeleton and the appendicular skeleton (Figure 41-5). The axial skeleton consists of 80 bones that make up the skull, vertebral column, and thorax. The appendicular skeleton consists of 126 bones that make up the upper and lower extremities, the shoulder girdle (pectoral girdle), and the pelvic girdle. The skeleton contributes about 14% of the weight of the adult body.
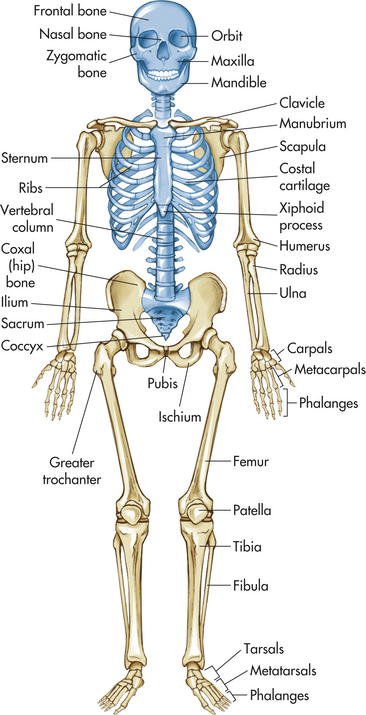
Figure 41-5 Anterior view of skeleton. Axial skeleton in blue; appendicular skeleton in tan. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St. Louis, 2010, Mosby.)
Bones can be classified by shape as long, flat, short (cuboidal), or irregular. Long bones are longer than they are wide and consist of a narrow tubular midportion (diaphysis) that merges into a broader neck (metaphysis) and a broad end (epiphysis) (see Figure 41-3).
The diaphysis consists of a shaft of thick, rigid compact bone that can tolerate bending forces. Contained within the diaphysis is the elongated marrow (medullary) cavity. The marrow cavity of the diaphysis contains primarily fatty tissue, which is referred to as yellow marrow. The yellow marrow assists red bone marrow in hematopoiesis only during times of stress. The yellow marrow cavity of the diaphysis is continuous with marrow cavities in the spongy bone of the metaphysis and diaphysis. The marrow contained within the epiphysis is red because it contains primarily blood-forming tissue (see Chapter 25). A layer of connective tissue, the endosteum, lines the outer surfaces of both types of marrow cavity.
The broadness of the epiphysis allows weightbearing to be distributed over a wide area. The epiphysis is made up of spongy bone covered by a very thin layer of compact bone. In a child the epiphysis is separated from the metaphysis by a cartilaginous growth plate, the epiphyseal plate. After puberty the epiphyseal plate calcifies and the epiphysis and metaphysis merge. By adulthood the line of demarcation between the epiphysis and metaphysis is undetectable.
In flat bones, such as the ribs or scapulae, two plates of compact bone are roughly parallel to each other. Between the compact bone plates is a layer of spongy bone. Short bones (cuboidal bones), such as the bones of the wrist or ankle, are often cuboidal in shape. They consist of spongy bone covered by a thin layer of compact bone.
Irregular bones, such as the vertebrae, mandibles, or other facial bones, have various shapes that include thin and thick segments. The thin part of an irregular bone consists of two plates of compact bone with spongy bone between the plates. The thick part consists of spongy bone surrounded by a layer of compact bone.
Maintenance of Bone Integrity
The internal structure of bone is maintained by remodeling, a three-phase process in which existing bone is resorbed and new bone is laid down to replace it. Remodeling is carried out by clusters of bone cells termed bone-remodeling units. The bone remodeling units are made up of bone precursor cells that differentiate into osteoclasts and osteoblasts. Precursor cells are located on the free surfaces of bones and along the vascular channels (especially the marrow cavities).
In phase 1 (activation) of the remodeling cycle, a stimulus (e.g., hormone, drug, vitamin, physical stressor) activates programmed osteocyte cell death (apoptosis). The distribution of these apoptotic osteocytes provides osteoclasts with information about where to begin resorbing damaged bone.12 In phase 2 (resorption), the osteoclasts form a “cutting cone,” which gradually resorbs bone, leaving behind an elongated cavity termed a resorption cavity. The resorption cavity in compact bone follows the longitudinal axis of the haversian system, and the resorption cavity in spongy bone parallels the surface of the trabeculae.
Phase 3 (formation) is the laying down of new bone, termed secondary bone, by osteoblasts lining the walls of the resorption cavity. Successive layers (lamellae) in compact bone are laid down until the resorption cavity is reduced to a narrow haversian canal around a blood vessel. In this way, old haversian systems are destroyed and new haversian systems are formed. New trabeculae are formed in spongy bone. The entire process of remodeling takes about 3 to 4 months.
Repair
The remodeling process can repair microscopic bone injuries, but gross injuries, such as fractures and surgical wounds (osteotomies), heal by the same stages as soft tissue injuries, except that new bone, instead of scar tissue, is the final result (see Chapter 6). In bone the stages of wound healing are as follows:
1. Hematoma formation: occurs if vessels have been damaged, causing hemorrhage. Fibrin and platelets within the hematoma form a meshwork that is the initial framework for healing with the help of hematopoietic growth factors such as platelet-derived growth factor and TGF-β (see Table 41-3).
2. Procallus formation: fibroblasts, capillary buds, and osteoblasts move into the wound to produce granulation tissue called procallus. Cartilage is formed as a precursor of bone, and types I, II, and III collagen are formed. Enzymes and growth factors, such as insulin and insulin-like growth factors, plus bone morphogenic protein and osteogenin, aid in this stage of healing.
3. Callus formation: osteoblasts in the procallus form membranous or woven bone (callus). Enzymes increase the phosphate content and permit the phosphate to join with calcium to be deposited as mineral to harden the callus.
4. Callus replacement: osteoblasts continue to replace the callus with lamellar bone or trabecular bone (Figure 41-6).
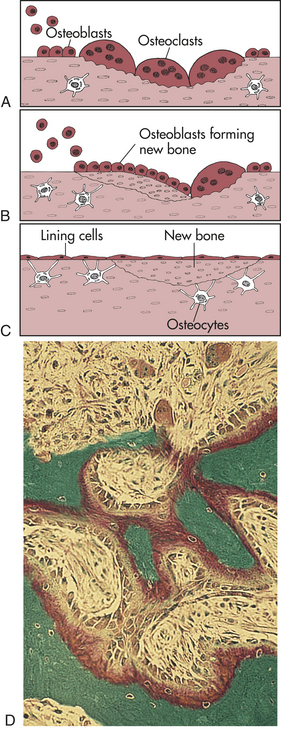
Figure 41-6 Bone remodeling. In the remodeling sequence, bone sections are removed by the bone-resorbing cells (osteoclasts) and replaced with a new section laid down by bone-forming cells (osteoblasts). The cells work in response to signals generated in that environment. The first phase of remodeling is mediated only by the multinucleated osteoclastic cells. They are activated, scoop out bone (A), and resorb it; then the work of the osteoblasts begins (B). Osteoblasts form new bone that replaces bone removed by the resorption process (C). The sequence takes 4 to 5 months. D, Active bone remodeling seen in the settings of primary or secondary hyperparathyroidism. Note the active osteoblasts surmounted on red-stained osteoid. Marrow fibrosis is present. (A to C from Mundy GR: Bone remodeling and its disorders, St Louis, 1995, Mosby. D from Damjanov I, Linder J, editors: Anderson’s pathology, ed 10, St Louis, 1996, Mosby.)
5. Remodeling: the periosteal and endosteal surfaces of the bone are remodeled to the size and shape of the bone before injury. Synthesis of other types of collagen recedes in favor of type I, which is the collagen found in bone. This final stage of healing, or remodeling, is vital because bone that has not been remodeled does not have good mechanical properties for weightbearing and mobility.
The speed with which bone heals depends on the severity of the bone disruption; the type and amount of bone tissue that must be replaced (spongy bone heals faster); blood supply and oxygen to the site; presence of growth and thyroid hormones, insulin, vitamins, and other nutrients; presence of systemic disease; effects of aging; and effective treatment, including immobilization and the prevention of complications such as infection. In general, however, hematoma formation occurs within hours of fracture or surgery; formation of procallus by osteoblasts occurs within days; callus formation occurs within weeks; and replacement and contour modeling occur within years—up to 4 years in some cases.
STRUCTURE AND FUNCTION OF JOINTS
The site where two or more bones meet is called a joint (articulation) (Figure 41-7). The primary function of joints is to provide stability and mobility to the skeleton. Whether a joint provides stability or mobility depends on its location and its structure. Generally, joints that stabilize the skeleton have a simpler structure than those that enable the skeleton to move. Most joints provide stability and mobility to some degree (Figure 41-8).
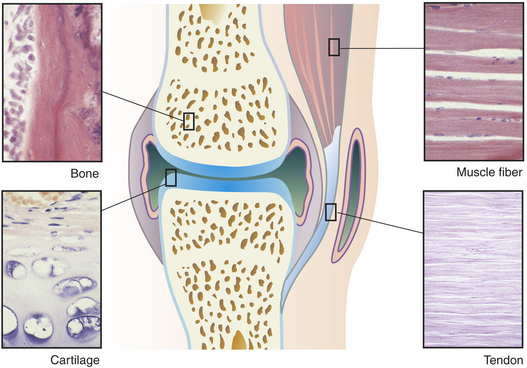
Figure 41-7 Main tissues of a joint. (Micrographs from Erlandsen SL, Magney JE: Color atlas of histology, St Louis, 1992, Mosby.)
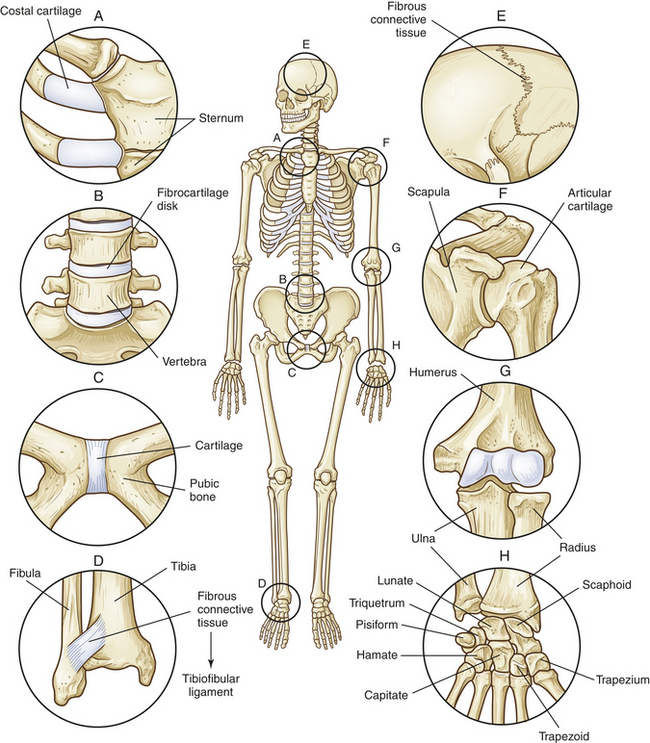
Figure 41-8 Types of joints. Cartilaginous (amphiarthrodial) joints, which are slightly movable, include (A) a synchondrosis that attaches ribs to costal cartilage; (B) a symphysis that connects vertebrae; and (C) the symphysis that connects the two pubic bones. Fibrous (synarthrodial) joints, which are immovable, include (D) the syndesmosis between the tibia and fibula; (E) sutures that connect the skull bones; and the gomphosis (not shown), which holds teeth in their sockets. The synovial joints include (F) the spheroid type at the shoulder; (G) the hinge type at the elbow; and (H) the gliding joints of the hand.
Joints are classified based on the degree of movement they permit or on the connecting tissues that hold them together. Based on movement, a joint is classified as a synarthrosis (immovable joint), an amphiarthrosis (slightly movable joint), or a diarthrosis (freely movable joint). On the basis of connective structures, joints are classified broadly as fibrous, cartilaginous, and synovial. Each of these three structural classifications can be subdivided according to the shape and contour of the articulating surfaces (ends) of the bones and the type of motion the joint permits.
Fibrous Joints
A joint in which bone is united directly to bone by fibrous connective tissue is called a fibrous joint. Generally, fibrous joints are synarthroses (immovable), but many fibrous joints allow some movement. The degree of movement depends on the distance between the bones and the flexibility of the fibrous connective tissue.
Fibrous joints are further subdivided into three types: sutures, syndesmoses, and gomphoses. A suture has a thin layer of dense fibrous tissue that binds together interlocking flat bones in the skulls of young children. Sutures form an extremely tight union that permits no motion. By adulthood the fibrous tissue has been replaced by bone. A syndesmosis is a joint in which the two bony surfaces are united by a ligament or membrane. The fibers of ligaments are flexible and stretch, permitting a limited amount of movement. The paired bones of the lower arm (radius and ulna) and the lower leg (tibia and fibula) and their ligaments are syndesmotic joints. A gomphosis is a special type of fibrous joint in which a conical projection fits into a complementary socket and is held there by a ligament. The teeth held in the maxilla or mandible are gomphosis joints.
Cartilaginous Joints
The two types of cartilaginous joints are symphyses and synchondroses. A symphysis is a cartilaginous joint in which bones are united by a pad or disk of fibrocartilage. The articulating surfaces of the two bones are usually covered by a thin layer of hyaline cartilage, and the thick pad of fibrocartilage acts as a shock absorber and stabilizer. Examples of symphyses are the symphysis pubis, which joins the two pubic bones, and the intervertebral disks, which join the bodies of the vertebrae. A synchondrosis is a joint in which hyaline cartilage, rather than fibrocartilage, connects the two bones. The joints between the ribs and the sternum are synchondroses. The hyaline cartilage of these joints is called costal cartilage. Slight movement at the synchondroses between the ribs and the sternum allows the chest to move outward and upward during breathing.
Synovial Joints
Synovial joints (diarthroses) are the most movable and the most complex joints in the body (Figure 41-9). A synovial joint consists of the following parts:
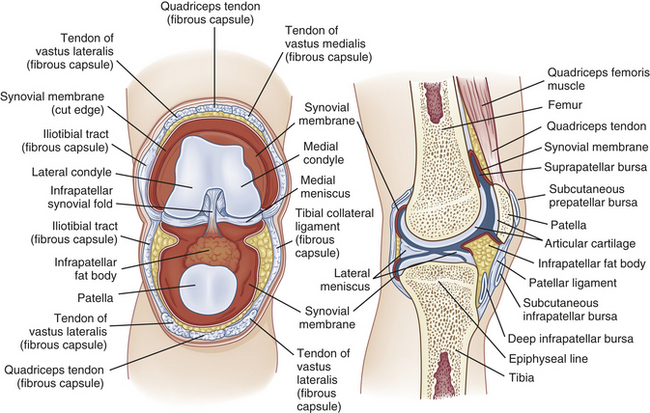
1. A fibrous joint capsule (articular capsule)
2. A synovial membrane that lines the inner surface of the joint capsule
3. A joint cavity (synovial cavity), a space formed by the capsule
4. Synovial fluid, which fills the joint cavity and lubricates the joint surface
5. Articular cartilage, which covers and pads the articulating bony surfaces
Joint Capsule: The fibrous joint capsule (articular capsule) is connective tissue that covers the ends of the bones where they meet in the joint. Sharpey fibers firmly attach the proximal and distal capsules to the periosteum and ligaments and tendons, which also reinforce the capsule. The joint capsule is made up of parallel, interlacing bundles of dense, white fibrous tissue. It is richly supplied with nerves, blood vessels, and lymphatic vessels. The nerves in and around the joint capsule are sensitive to the rate and direction of motion, compression, tension, vibration, and pain.
Synovial Membrane: The synovial membrane (synovium) is the smooth, delicate inner lining of the joint capsule (Figure 41-10). It lines the nonarticular portion of the synovial joint and any ligaments or tendons that traverse the joint cavity. It is made up of two layers—a vascular layer (subintima) and a thin cellular layer (intima). The vascular subintima merges with the fibrous joint capsule and is composed of loose fibrous connective tissue, elastin fibers, fat cells, fibroblasts, macrophages, and mast cells. The intima consists of rows of synovial cells embedded in a fiber-free intercellular matrix. The intima contains two types of synovial cells: type A and type B. Type A synovial cells ingest and remove bacteria and particles of debris by phagocytosis in the joint cavity. (Phagocytosis is described in Chapter 6.) Type B synovial cells secrete hyaluronate, a binding agent that gives synovial fluid its viscous quality. The synovial membrane is richly supplied with blood and lymphatic vessels; therefore, it is capable of rapid repair and regeneration.
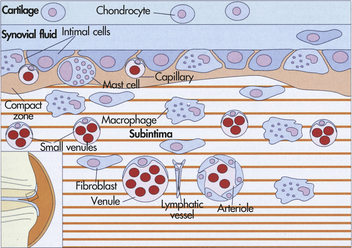
Joint Cavity: The joint cavity (synovial cavity) is an enclosed fluid-filled space between the articulating surfaces of the two bones. This small cavity, often called the joint space, enables the two bones to move “against” one another. The synovial cavity is surrounded by the synovial membrane and filled with a clear, viscous, slick fluid called the synovial fluid.
Synovial Fluid: Synovial fluid is superfiltrated plasma from blood vessels in the synovial membrane. Synovial fluid lubricates the joint surfaces, nourishes the pad of the articular cartilage that covers the ends of the bones, and contains free-floating synovial cells and various leukocytes that phagocytose joint debris and microorganisms. Loss of synovial fluid leads to rapid deterioration of articular cartilage.
Articular Cartilage: Articular cartilage is a layer of hyaline cartilage that covers the end of each bone (Figure 41-11). It ranges from 2 to 5 mm thick, depending on the size of the joint, the fit of the two bone ends, and the amount of weight and shearing force the joint normally withstands. Cartilage strength and its biologic properties are due to an extensive network of cross-linked collagen fibers.13 Figure 41-12 illustrates how collagen cross-links from cartilage. The function of articular cartilage is to reduce friction in the joint and to distribute the forces of weightbearing. Articular cartilage is composed of chondrocytes (cartilage cells) (making up about 2% of the tissue) and an intercellular matrix made up of collagen (making up about 10% to 30% of weight), protein polysaccharides (making up 5% to 10% of weight), and water. The water content ranges from 60% to almost 80% of the net weight of the cartilage, and individual molecules rapidly enter or exit the articular cartilage to contribute to the resiliency of the tissue.
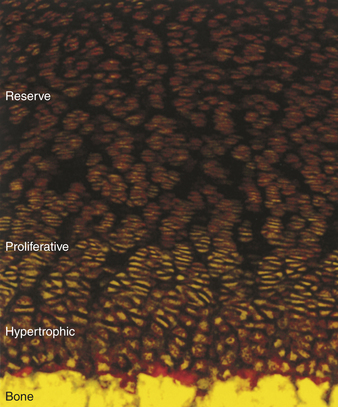
Figure 41-11 Collagen zones. The three collagen zones (reserve, proliferative, and hypertrophic) are distinctly shown in a growth plate. (From Hjorten R et al: Bone 41[4]:535, 2007.)
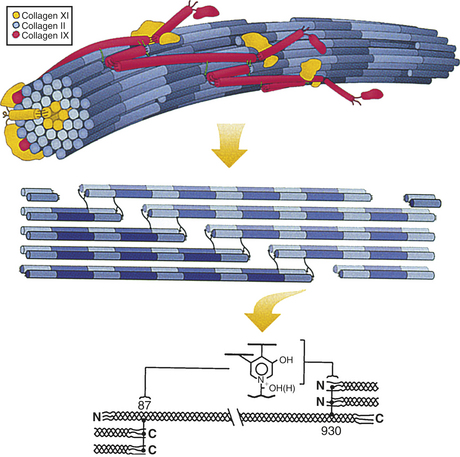
Figure 41-12 Heterotypic collagen fibril. Hierarchical depiction of heterotypic collagen fibril, emphasizing the internal axial relationships required for mature cross-link formation. Upper drawing: Three-dimensional concept of the type II/IX/XI heterotypic fibril of developing cartilage matrix. Middle drawing: Detail illustrating required nearest neighbor axial relationships for trifunctional intermolecular cross-links to form in collagens of cartilage, bone, and other high-tensile strength tissue matrices. The exact 3D spatial pattern of cross-linking bonds is still unclear for any tissue. Lower drawing: Detail of the axial stagger of individual collagen molecules required for pyridinoline cross-linking. (From Eyre DA, Weiss MA, Wu JJ: Methods 4[1]:65, 2008.)
The intercellular matrix is produced by the chondrocytes, which synthesize and extrude collagen that, like the collagen produced by bone cells, is distributed throughout the cartilage in a highly organized system of fibers. Collagen fibers in cartilage are made up of many fine fibrils that, like bone fibrils, are assembled in an orderly fashion that makes them resistant to physical, metabolic, or chemical breakdown. The main differences between bone collagen and cartilage collagen are the amino acid content of the α-chains and the composition of the fibrils. Bone collagen fibrils are made up of two type I chains and one type II chain. Approximately 90% of the cartilage collagen fibrils is made up of three identical type II chains, with the remaining 10% made up of types V, VI, IX, X, and XI.
At the surface of articular cartilage, the collagen fibers run parallel to the joint surface and are closely compacted into a dense, protective mat. (Loss of this dense, compacted configuration at the surface subjects the underlying fibers to splitting and thinning, in which case the cartilage is unable to tolerate weightbearing.) In the middle layer (the proliferative zone) of the cartilage, the fibers are arranged tangential to the surface, which allows them to deform and absorb some of the force of weightbearing. In the bottom layer (the hypertrophic zone) of the cartilage, the fibers are perpendicular to the joint surface, allowing them to resist shear forces, and are embedded in a calcified layer of cartilage called the tidemark. The tidemark anchors the collagen fibers to the underlying (subchondral) bone and represents the zone between calcified and uncalcified cartilage. Collagen fibers are important components of the cartilage matrix because they account for approximately 60% of the dry weight and because they (1) anchor the cartilage securely to underlying bone, (2) provide a taut framework for the cartilage, (3) control the loss of fluid from the cartilage, and (4) prevent the escape of protein polysaccharides (proteoglycans) from the cartilage.
The proteoglycans are macromolecules consisting of proteins, carbohydrates (glycosaminoglycans), and hyaluronic acid. The glycosaminoglycans (keratan sulfate and chondroitin sulfate) are attached to the protein core, and several protein cores (with their attached glycosaminoglycans) are bound to a hyaluronic acid chain by a special protein called link protein. The proteoglycans give articular cartilage its stiff quality and regulate the movement of synovial fluid through the cartilage. Without proteoglycans, normal weightbearing would rapidly and completely press all the synovial fluid out of the cartilage. The proteoglycans act as a pump, permitting enough fluid to be pressed out to ensure that a fluid film is always present on the surface of the cartilage, even after hours of weightbearing. The pumping action of proteoglycans also draws synovial fluid back into the cartilage after a weightbearing load is released. Mobility and weightbearing are necessary for the pumping action of proteoglycans to occur. Nonuse of a joint quickly reduces the pumping action, which changes the composition of the matrix and interferes with the nutrition of the chondrocytes.
Articular cartilage has no blood vessels, lymph vessels, or nerves. Therefore, it is insensitive to pain and regenerates slowly and minimally after injury. Regeneration occurs primarily at sites where the articular cartilage meets the synovial membrane, where blood vessels and nutrients are available. In general, it has been difficult to enhance cartilage repair, but that may be changing (see What’s New? Progress in Rebuilding Cartilage).
Movement
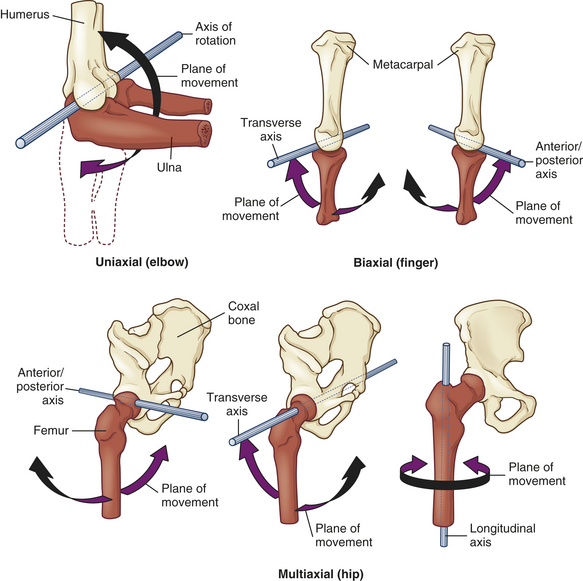
Synovial joints are described as uniaxial, biaxial, or multiaxial according to the shapes of the bone ends and the type of movement occurring at the joint (Figure 41-13). Usually one of the bones is stable and serves as an axis for the motion of the other bone. The body movements made possible by various synovial joints are either circular or angular (Figure 41-14).
STRUCTURE AND FUNCTION OF SKELETAL MUSCLES
The skeletal muscles are made up of millions of individual fibers that by the process of contraction and relaxation do the work necessary to complete movements as varied as a ballerina’s pirouette or an artist’s deft stroke (Figure 41-15). Muscle constitutes 40% of adults’ body weight and 50% of children’s body weight. Muscle is 75% water, 20% protein, and 5% organic and inorganic compounds; 32% of all protein stores for energy and metabolism is contained in muscle.
WHAT’S NEW?
Progress in Rebuilding Cartilage
Efforts to repair damaged cartilage have been difficult because of cartilage’s poor vascularity. Currently available techniques, such as autologous chondrocyte transplantation, have not consistently produced good results. Developments in tissue engineering have been aided by maintaining improved biologic control of cell growth, differentiation, and metabolic activity.
Techniques known as “functional tissue engineering” should not only provide a network of chondrocyte cell growth but also should greatly improve the biomechanical properties of engineered cartilage. By more closely resembling native cartilage, these technologies show great promise for clinically improved outcomes in cartilage repair and replacement.
Data from Guilaf F: Am Acad Orthop Surg 16(1):58, 2008.
Whole Muscle
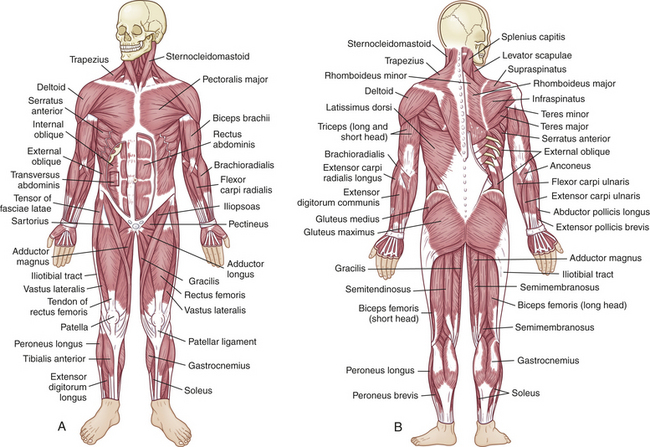
There are more than 350 named muscles; almost all are paired. The body’s muscles vary dramatically; they range from 2 to 60 cm in length and are shaped according to function. Fusiform muscles are elongated muscles shaped like straps and can run from one joint to another. Pennate muscles are broad, flat, and slightly fan shaped, with fibers running obliquely to the muscle’s long axis. The multipennate deltoid muscle, which flexes and extends the arm, is a good example of a muscle shaped according to its function.
Each skeletal muscle is a separate organ encased in a three-part connective tissue framework called fascia. The layers of connective tissue protect the muscle fibers, attach the muscle to bony prominences, and provide a structure for a network of nerve fibers, blood vessels, and lymphatic channels.
The outermost layer, the epimysium, is located on the surface of the muscle and tapers at each end to form the tendon (Figure 41-16). Tendons allow a short muscle to exert power on a distant joint, whereas a thick muscle interferes with joint mobility. The next layer, the perimysium, further subdivides the muscle fibers into bundles of connective tissue, or fascicles. The endomysium surrounds the muscle fascicles, the smallest unit of muscle fibers visible without a microscope. The ligaments, tendons, and fascia are made up of connective tissue that also serves to buffer the limbs from the effects of sudden strains or changes in speed. The rapid recovery necessary for strenuous exercise is supported by the elastic property of muscle and its connective tissue.
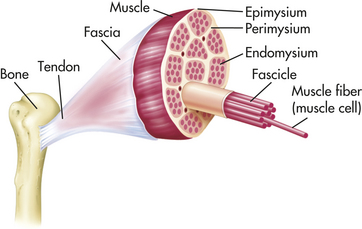
Figure 41-16 Cross section of skeletal muscle showing muscle fibers and their coverings. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 5, St Louis, 2003, Mosby.)
Skeletal muscle is described, almost interchangeably, as voluntary, striated, or extrafusal. “Voluntary” indicates that the muscle is controlled directly by the central nervous system. “Striated” describes the striated, or striped, pattern of skeletal muscle viewed under a light microscope. The striations result from the organization of the muscle fibers into the contractile units called sarcomeres. “Extrafusal” distinguishes the skeletal muscle fibers from other contractile fibers located within the sensory organs of the muscle.
Other components that are visible on gross inspection of the whole muscle include the motor and sensory nerve fibers. These function together with the muscle, innervating portions of it and providing the electrical impulses needed for motor function.
Motor Unit
From the anterior horn cell of the spinal cord, the axons of motor nerves branch out to innervate a specific group of muscle fibers. Each anterior horn cell, its axon (part of a lower motor neuron; see Chapter 14), and the muscle fibers innervated by it are called a motor unit (Figure 41-17). The motor units are composed of lower motor neurons, which extend to skeletal muscles. Often termed the functional unit of the neuromuscular system, the motor unit behaves as a single entity and contracts as a whole when it receives an electrical impulse.
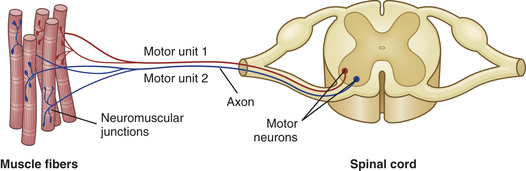
Figure 41-17 Motor units of a muscle. Each motor unit consists of a somatic motor neuron and all the muscle fibers (cells) supplied by the neuron and its axon branches.
The whole muscle may be controlled by several motor nerve axons. These branch to innervate many motor units within the muscle. The whole muscle then may be made up of many motor units. The number of motor units per individual muscle varies greatly. In the calf, for example, one motor axon will innervate approximately 2000 muscle fibers, out of a total of 1,200,000 muscle fibers. This is a high innervation ratio of muscle fibers to axons, and it contrasts markedly with the low innervation ratio in the laryngeal muscles. There, two or three muscle fibers constitute each motor unit, and the innervation ratio can be of great functional significance. The greater the innervation ratio of a particular organ, the greater its endurance. Higher innervation ratios prevent fatigue, and lower innervation ratios allow for precision of movement.
Sensory Receptors: Although muscles function as effector organs, they also contain sensory receptors and are involved in sending different signals to the central nervous system. Among these are the muscle spindles and Golgi tendon organs. Spindles are mechanoreceptors that lie parallel to muscle fibers and respond to muscle stretching. Golgi tendon organs are dendrites that terminate and branch to tendons near the neuromuscular junction. Motor and sensory neurons secrete a proteoglycan called neuroregulin (NRG) that increases acetylcholine receptors and helps in the formation of muscle spindle fibers.14 The muscle spindles, Golgi tendon organs, and free nerve endings provide a means of reporting changes in length, tension, velocity, and tone in the muscle. This system of afferent signals is responsible for the muscle stretch response and maintenance of normal muscle tone.
Muscle Fibers: Each muscle fiber is a single muscle cell. This long cell is cylindrical and surrounded by a membrane capable of excitation and impulse propagation. The muscle fiber contains bundles of myofibrils, the fiber’s functional subunits, in a parallel arrangement along the longitudinal axis of the muscle (Figure 41-18). At birth the muscle fibers have completed development from precursor cells called myoblasts. All voluntary muscles are derived from the mesodermal layer of the embryo.
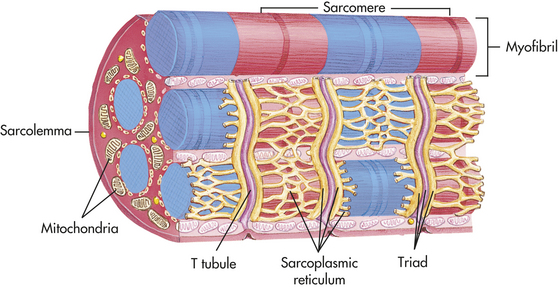
Figure 41-18 Myofibrils of a skeletal muscle fiber (cell) and overall organization of skeletal muscle. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 5, St Louis, 2003, Mosby.)
The type of peripheral nerve influences the muscle fiber and motor unit considerably. Whether motor nerves are fast or slow determines the type of muscle fibers in the motor unit. Type II fibers, also called white fast-motor fibers, are innervated by relatively large type II alpha motor neurons with fast conduction velocities. These fibers rely on a short-term anaerobic glycolytic system for rapid energy transfer, whereas type I fibers depend on aerobic oxidative metabolism. Histochemical stains are now routinely used to describe the structure of muscle fibers and contractile elements of muscle biopsy specimens. White muscle (type II fibers) stains dark in the enzyme stain adenosine triphosphatase (ATPase) at a pH of 9.4. Red muscle (type I fibers) appears lightly stained.
The overlap of muscle fibers that appear with staining gives the checkerboard appearance of muscle biopsy specimens and provides an equal distribution of fiber types throughout the muscle. This overlap also helps compensate for muscle fiber loss and fatigue of individual motor units during activity. In spite of this, some muscles contain proportionally more of one fiber type than another. The postural muscles have more type I fibers, allowing them the high resistance to fatigue that is necessary to maintain the same position for extended periods. The ocular muscles have more type II muscle fibers, allowing them to respond rapidly to visual changes. (Table 41-6 describes the specific characteristics of type I and type II fibers.)
Table 41-6
Characteristics of Muscle Fibers
| Characteristic | Type I (Red) | Type II (White) |
| Anatomic location | Deep axial portion of surface muscle | Surface portion of surface muscle |
| Contraction speed | Slow | Fast |
| Motor neuron type | Type I, small alpha | Type II, large alpha |
| Firing frequency | Low, long duration | Rapid, short duration |
| Resistance to fatigue | High | Low |
| Myoglobin | High | Low |
| Capillary supply | Profuse | Intermediate to sparse |
| Metabolism | Oxidative | Glycolysis |
| Mitochondria | Many | Few |
| Enzymes | Lactate dehydrogenase, types 1 to 3 | Lactate dehydrogenase, types 4 and 5 |
| Creatine kinase | Cardiac type | Fast, skeletal |
| Example (most muscles are mixed) | Greater proportion of slow-contracting fibers in soleus | Greater proportion of fast-contracting fibers in laryngeal and ocular muscles |
| Glycogen content | Low | High |
| Intensity of contraction | Low | High |
| Aerobic metabolic capacity | High | Low |
| Fiber diameter | Small | Large |
| Myosin-adenosine triphosphatase (ATPase) activity | Low | High |
From Spence AP, Mason EE: Human anatomy and physiology, ed 4, St Paul, MN, 1992, West Publishing.
The number of muscle fibers varies according to location. Large muscles, such as the gastrocnemius, have more fibers (1,200,000) than the smaller muscles, such as the lumbrical muscles in the hand (10,000). The diameter of muscle fibers also varies. The closely packed polygons are small (10 to 20 μm) until puberty, when they attain the normal adult diameter of 40 to 80 μm. Women usually have smaller-diameter fibers than men. Small muscles, such as the ocular muscles, are 15 μm in diameter; larger, more proximal muscles are 40 μm. Fiber size can have functional significance. Studies have shown that larger fiber diameter is associated with generation of greater forces. Fiber diameter can be increased by activities that cause hypertrophied muscle, such as exercise or occupational overuse.
The major components of the muscle fiber include the muscle membrane, myofibrils, sarcotubular system, sarcoplasm, and mitochondria (see Figure 41-18). The muscle membrane is a two-part membrane. It includes the sarcolemma, which contains the plasma membrane of the muscle cell, and the cell’s basement membrane. The sarcolemma is 7.5 nm thick and is capable of propagating electrical impulses to initiate contraction. At the motor nerve end plate, where the nerve impulse is transmitted, the sarcolemma forms the highly convoluted synaptic cleft. The sarcolemma is made up of lipid molecules and protein systems. The protein systems perform special functions, such as transport of nutrients and protein synthesis. They also provide the sodium-potassium pump and include the cell’s cholinergic receptor. The basement membrane is 50 nm thick and is composed primarily of proteins and polysaccharides. It serves as the cell’s microskeleton and maintains the shape of the muscle cell. The basement membrane also may function in some way to restrict further diffusion of electrolytes once they have crossed the sarcolemma.
The sarcoplasm is the cytoplasm of the muscle cell and contains the intracellular components that are common to all cells (see Chapter 1). The sarcoplasm is an aqueous substance that provides a matrix that surrounds the myofibrils. It contains numerous enzymes and proteins that are responsible for the cell’s energy production, protein synthesis, and oxygen storage. The mitochondria house enzyme systems for energy production, particularly those that regulate such processes as the citric acid cycle and adenosine triphosphate (ATP) formation. Many other structures are present in the sarcoplasm. The ribosomes are composed primarily of ribonucleic acid (RNA) and participate in the process of protein synthesis. The cell nucleus, satellite cells, glycogen granules, and lipid droplets are suspended in the sarcoplasmic matrix. Blood vessels, nerve endings, muscle spindles, and Golgi tendon organs are also directly located within this structure.
Unique to the muscle is the sarcotubular system, a network that includes the transverse tubules and the sarcoplasmic reticulum, which crosses the interior of the cell. The sarcoplasmic reticulum is made in the same manner as the endoplasmic reticulum in other cells. In the muscle cells the sarcoplasmic reticulum is involved in calcium transport, which initiates muscle contraction at the sarcomere, a portion of the myofibril. The sarcoplasmic reticulum is composed of tubules that run parallel to the myofibrils. The longitudinal tubules are termed sarcotubules. The transverse tubules, which are closely associated with the sarcotubules, run across the sarcoplasm and communicate with the extracellular space. Together, the tubules of this membrane system allow for intracellular calcium uptake, regulation, release during muscle contraction, and storage of calcium during muscle relaxation.
Myofibrils: The myofibrils are the functional units of muscle contraction. Each myofibril contains sarcomeres, which appear at intervals (see Figure 41-18). The sarcomeres are composed of two contractile proteins, actin and myosin. The sarcomere is responsible for converting chemical energy into movement.
The myofibrils are the most abundant subcellular muscle component, equaling 85% to 90% of the total volume. On cross section they are irregular polygons with a mean diameter of less than 1 μm. Each myofibril is composed of serially repeating sarcomeres, separated by Z lines that give the muscle its striped, cross-striated appearance. Each sarcomere has a dark A band and is flanked by two light I bands (Figure 41-19). Within the sarcomere is the giant muscle protein, titin. Titin functions as a molecular spring that plays a significant role in determining muscle stiffness and elasticity.15 The A band is 1.5 to 1.6 μm long and contains thick myosin filaments. Included in the A band is a lighter zone called the H band, and in the center of the H band is the dark M band, or M line. The I band, which contains actin, is divided at the midpoint of each sarcomere by the Z line. Its length varies with the start of muscle contraction.
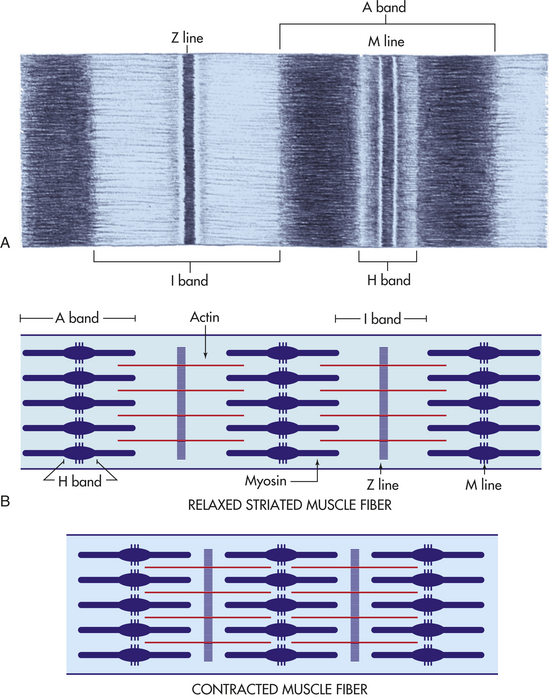
Figure 41-19 Muscle fibers. A, Lines and bands in striated muscle. B, Relationships of bands, actin, myosin, and lines in relaxed and contracted muscle fibers. (A modified from Thompson JM et al: Mosby’s clinical nursing, ed 5, St Louis, 2002, Mosby.)
Myofibrils are composed of myofilaments. Each myofilament is structured in a closely packed hexagonal arrangement, with two thin filaments for every thick filament. The thick filament, along with C protein and M line protein, is made up of myosin. Myosin has two subunits, heavy and light meromyosin, which resemble twisted golf club shafts. The thin filaments are twisted double strands made up of actin, troponin, and tropomyosin (see Chapter 29 and Figure 28-15).
Muscle Proteins: Table 41-7 summarizes muscle protein distribution, location, and possible functional significance. The contractile and regulatory functions of actin, myosin, and the troponin-tropomyosin complex (associated with actin) are the most commonly known. They also account for most of the protein found in the myofibril. The structural and regulatory processes of muscle proteins are less well understood. Alpha actin and beta actin are known to link the filaments. M protein contains the enzyme creatine kinase (CK). Creatine is released when muscle cells are damaged, making serum creatine an important test of pathologic conditions of muscles.
Table 41-7
Contractile Proteins of Skeletal Muscle Fibrils
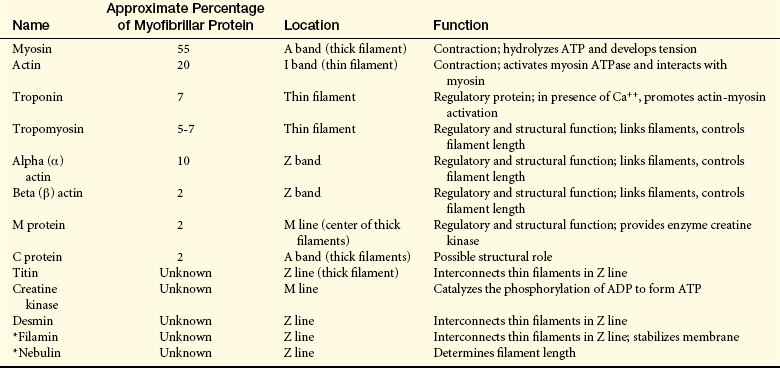
ADP, adenosine diphosphate; ATP, Adenosine triphosphate; ATPase, adenosine triphosphatase.
∗Data from Ma K, Wang K: Fed Eur Biochem Soc Lett 532(3):273-278, 2002; Sampson LJ, Leyland ML, Dart D: J Biol Chem 278(43):41988-41997, 2003.
Modified from Simon SR, editor: Orthopaedic basic science, Chicago, 1994, American Academy of Orthopaedic Surgeons.
The most abundant proteins, actin and myosin, are also found in other cells, particularly motile cells such as platelets. The complete amino acid sequences of actin and myosin have been identified. Noteworthy is the presence of the amino 3-methylhistidine, found only in the thin filament, actin; 85% to 90% of 3-methylhistidine is found in skeletal muscle. Because it is excreted unchanged (in the urine) after release from muscle and other tissue, 3-methylhistidine has been used to gauge muscle protein degradation. The amino acids lysine and histidine, in addition to leucine, have been used to study protein synthesis by means of stable isotope infusion and muscle biopsy analysis.
Nonprotein Constituents of Muscle: Nitrogen, creatine, creatinine, phosphocreatine, purines, uric acid, and amino acids serve in the complex process of muscle metabolism. Phosphocreatine concentration is an extremely sensitive indicator of muscle fiber activity.16 Glycogen and its derivatives are present as energy sources.
Creatine and creatinine metabolism have been used to measure muscle mass. Plasma creatine is taken up by muscle and converted into the high-energy phosphate compound phosphocreatine by the enzyme creatine kinase. Creatinine is formed in muscle from creatine at a constant rate of 2% per day. (Tests for plasma creatine are discussed in Chapter 35.) Creatine excretion is increased in muscle wasting. This change reflects the reduction in total body creatine stores and loss of muscle mass.
Inorganic compounds, anions (phosphate, chloride), and cations (calcium, magnesium, sodium, potassium) are important in regulating protein synthesis, muscle contraction, enzyme systems, and membrane stabilization. Total body potassium (TBK), measured by the K40 method, has been used to measure muscle mass, also called lean body mass. TBK levels reflect changes in muscle mass seen during growth, malnutrition, and muscle wasting.
Components of Muscle Function
The ultimate function of muscle is to accomplish work. Although variously expressed in such measures as foot-pounds or kilogram-meters, work usually refers to the amount of energy liberated or force exerted over a distance (work = force × distance). Muscles usually contract or tense while doing work. Muscle contraction occurs on the molecular level and leads to the observable phenomenon of muscle movement.
Muscle Contraction at the Molecular Level
Muscle contraction is a four-step process: excitation, coupling, contraction, and relaxation. The initial contraction process is the excitation-contraction coupling (ECC) series, which involves the electrical properties of all cells and the movement of ions across the plasma membrane (see Chapter 1). The muscle fiber is an excitable tissue. At rest an electric charge of −90 mV is continually maintained across the sarcolemma. This resting potential, generated by the separation of positive and negative charges on either side of the membrane, creates an electrochemical equilibrium caused by the selective permeability of the sarcolemma to electrolytes in the intracellular and extracellular fluids, particularly potassium and sodium.
Excitation, the first step of muscle contraction, begins with the spread of an action potential from the nerve terminal to the neuromuscular junction. The rapid depolarization of the membrane initiates an electrical impulse in the muscle fiber membrane called the muscle fiber action potential. As the action potential advances along the sarcolemmal membrane, it spreads to the transverse tubules. (The velocity of conduction is much slower in muscle fibers than in myelinated nerve fibers—only 3 to 5 m/second compared with 54 to 90 m/second in nerve fibers.)
The second stage, coupling, follows the depolarization of the transverse tubules. This stage consists of the migration of calcium ions, which are stored in the sarcoplasmic reticulum, to the myofilaments. Calcium affects troponin and tropomyosin, muscle proteins that bind with actin when the muscle is at rest. In the presence of calcium, however, both of these proteins are attracted to calcium ions, leaving the actin free to bind with myosin.
Contraction begins as the calcium ions combine with troponin, a reaction that overcomes the inhibitory function of the troponin-tropomyosin system. The thin filament actin then slides toward the thick filament, myosin. The two ends of the myofibril shorten after contraction when the myosin heads attach to the actin molecules, forming a cross-bridge that constitutes an actin-myosin complex. ATP, located on the actin-myosin complex, is released when the cross-bridges attach. This is the sliding filament theory described by A.F. Huxley in the 1950s, but it is now called the cross-bridge theory because of the formation of the actin-myosin cross-bridges. The process is so named because the actin actually slides onto the myosin, causing the sarcomere to shorten. The useful distance of contraction of a skeletal muscle is approximately 25% to 35% of the muscle’s length.
The last step, relaxation, begins as the sarcoplasmic reticulum absorbs the calcium molecules, removing them from interaction with troponin. Calcium is pumped back into the sarcoplasmic reticulum by means of an active transport process. The cross-bridges detach, and the sarcomere lengthens. (The cross-bridge theory of muscle contraction is discussed in Chapter 29.)
Muscle Metabolism
Skeletal muscle requires a constant supply of ATP and phosphocreatine. These substances are necessary to fuel the complex processes of muscle contraction, driving the cross-bridges of actin and myosin together and transporting calcium from the sarcoplasmic reticulum to the myofibril. Other internal processes of the muscular system that require ATP include protein synthesis, which replenishes muscle constituents and accommodates growth and repair. The rate of protein synthesis is related to hormone levels (particularly insulin), amino acid substrates, and overall nutritional status. At rest the rate of ATP formation by oxidation of glucose or acetoacetate is sufficient to maintain internal processes, given normal nutritional status. During activity the need for ATP increases 100-fold. The metabolic pathways for muscle activity in Table 41-8 show reactions to the immediate need for increased ATP caused by contraction. Activity lasting longer than 5 seconds expends the available stored ATP and phosphocreatine.
Table 41-8
Energy Sources for Muscular Activity

From Spence AP, Mason EE: Human anatomy and physiology, ed 4, St Paul, MN, 1992, West Publishing.
Stored glycogen and blood glucose are converted anaerobically to sustain brief activity without increasing the demand for oxygen. Anaerobic glycolysis is much less efficient than aerobic glycolysis, using six to eight times more glycogen to produce the same amount of ATP. With increased activity, such as intense exercise, or ischemia, an increase in lactic acid occurs because of the breakdown of glycogen, thus causing a shift in muscle pH (see Table 41-8). This short-term mechanism “buys time” by allowing ATP formation in spite of inadequate energy stores or oxygen supply. When the anaerobic threshold is reached and more oxygen is required, physiologic changes occur, including an increase in lactic acid and increases in oxygen consumption, heart rate, respiratory rate, and muscle blood flow.
Strenuous exercise requires oxygen, which activates the aerobic glycogen pathway for ATP formation. During maximal exercise, free fatty acid mobilization and the aerobic glycogen pathways provide ATP over an extended time. These pathways require oxygen to maintain maximal activity and return the muscle to the resting state. Maximal exercise increases oxygen uptake 15 to 30 times over the resting state.17 When this system becomes exhausted or inadequate to respond to the need for ATP, fatigue and weakness finally force the muscle to reduce activity, with a resultant buildup of lactic acid in muscle fibers.
The ability to sustain maximal muscular activity leads to the accumulation of oxygen debt. Oxygen debt is the amount of oxygen needed to convert the buildup of lactic acid to glucose and replenish ATP and phosphocreatine stores. For example, after running at maximal speed for 10 seconds, the average person has consumed 1 L of oxygen. At rest, oxygen consumption for the same period is approximately 40 ml. As the person recovers, the measured oxygen debt is 4 L greater than the amount used during activity.
Oxygen consumption is measured to calculate the metabolic cost of activity in normal and diseased muscle. It is an indirect measure of energy expenditure, along with timed tests of activity, heart rate, and respiratory quotient (ratio of carbon dioxide to expired oxygen consumed). Energy expenditure is measured directly by heat production because heat is released whenever work is accomplished.
Another factor that changes energy requirements is muscle fiber type. Type II fibers rely on anaerobic glycolytic metabolism and fatigue readily. Type I fibers can resist fatigue for longer periods because of their capacity for oxidative metabolism.
Muscle Mechanics
Muscle contraction cannot be viewed in isolation. Several factors determine how force is transmitted from the cross-bridges on individual muscle fibers to accomplish whole-muscle contraction. First, when a motor unit responds to a single nerve stimulus, it develops a phasic contraction, also called twitch. Because the motor unit contracts in an “all or nothing” manner, the contraction that is generated will be a maximal contraction. The central nervous system smoothly grades the force generated by “recruiting” additional motor units and varying the discharge frequency of each active motor unit. This adding of motor units within the muscle is called repetitive discharge.
Recruitment and repetitive discharge of motor units allow the muscle to activate the number of motor units needed to generate the desired force. The total force developed is the sum of the force generated by each motor unit. As the strength, speed, and duration of stimuli increase, the summation of contractions reaches a critical frequency called tetanus. When tetanus is reached, no further increase in force can be achieved.
Other variables, such as fiber type, innervation ratio, muscle temperature, and muscle shape, influence the efficiency of muscular contraction. The two muscle fiber types differ in their responses to electrical activity. Tetanus and duration of phasic contractions, which take microseconds to accomplish, are achieved more rapidly in type II than in type I muscle fibers. Low-innervation ratios promote control and coordination, whereas high ratios promote strength and endurance. Muscles work best at normal body temperature, 37° C (98.6° F). Finally, muscles with a large cross-sectional area, such as the fan-shaped pennate muscles, develop greater contractile forces than smaller-diameter muscles. The initial length of a muscle and the range of shortening that occur when the muscle contracts also determine the forces it can generate. The long fusiform muscles have a greater range of shortening and can contract up to 57% of their resting length. A certain amount of elongation is necessary to generate sufficient tension and muscular force. The elongation that occurs during the swing of a golf club or tennis racquet is an example of how stretch improves contractile force.
Types of Muscle Contraction
During isometric contraction (static or holding contraction), the muscle maintains constant length as tension is increased. Isometric contraction occurs, for example, when the arm or leg is pushed against an immovable object. The muscle contracts, but the limb does not move.
During isotonic contraction (lengthening or shortening contraction), the muscle maintains a constant tension as it moves. The terms “eccentric” for lengthening and “concentric” for shortening are technically inaccurate descriptions of muscle movement; the terms “lengthening” and “shortening” are more precise. Positive work is accomplished during shortening, and energy is released to exert force or lift a weight. In contrast, during lengthening the muscle lengthens and absorbs energy. Negative work is accomplished on the muscle by the load. Lengthening requires less energy to accomplish and may result in the development of pain and stiffness after unaccustomed exercise.
Movement of Muscle Groups
Muscles do not act alone but rather in groups, often under automatic control. When a muscle contracts and acts as a “prime mover,” or agonist, its reciprocal muscle, or antagonist, relaxes. This is easily tested by holding the right arm in the horizontal position in front of the body and then bending the elbow while feeling the upper arm, biceps in the front and the triceps in the back, with the other hand. The biceps is firm, and the triceps is soft. As the arm is extended, the muscles change. When the elbow is completely extended, the biceps is soft and the triceps firm. Completing this movement causes the agonist and antagonist to change automatically; only the movement is commanded, not the alternate contraction and relaxation of the specific muscle groups.
Other associated actions may be seen during walking; as the foot leaves the ground, the paravertebral and gluteal muscles on the opposite sides of the body contract to maintain balance. One notices the loss of the associated muscle’s action when paralysis offsets this process and decreases balance. If a person is paralyzed, difficulty in maintaining balance is noticeable.
TESTS OF MUSCULOSKELETAL FUNCTION
Diagnostic procedures to evaluate bone function include gait analysis, serum calcium and phosphorus, and imaging studies. Most imaging techniques provide morphologic rather than functional information about bone. Roentgenograms visualize bone structure, because bone absorbs x-ray beams better than soft tissue. Magnetic resonance imaging (MRI) is useful for evaluating primary or metastatic bone lesions, infection, marrow edema, bony erosions, osteonecrosis, fractures, and other pathologic changes of bone. Bone scanning is used to evaluate bone metabolism. After a small amount of radioactive tracer is injected, a special camera is used to identify bone absorption of the tracer. Single- or dual-photon absorptiometry is often used to measure density of bones in the extremities (single-photon absorptiometry) and fracture risk of vertebral bodies and the femoral neck (dual-photon absorptiometry). Dual-photon absorptiometry allows the soft tissue component to be subtracted. New technology promises more accurate evaluation of bone (see What’s New? Quantitative Assessment of Macro- and Microstructural Features of Bone).
Bone resorption is evaluated with urinary and serum measurements of cross-linked N-terminal telopeptides (NTx), a product of osteoclast bone resorption. NTx is specific for bone because the cross-links assessed are characteristic of bone collagen alone. Urine NTx is a more sensitive and specific biochemical marker of bone resorption than serum Ntx.18,19
Tests of Joint Function
Procedures used to diagnose joint function include arthrography, arthroscopy, magnetic resonance imaging, and synovial fluid analysis. Arthrography (the injection of dye into the joint) is particularly useful to diagnose tears in the fibrocartilage of the knee (meniscus) and the rotator cuff of the shoulder. Arthroscopy is the direct visualization of a joint through an arthroscope. Magnetic resonance imaging (MRI) produces images of body tissues through the use of electromagnetic (radio) waves that alter the atoms (hydrogen ions) in the nuclei of cells being examined. When the polarized radio waves are stopped, the nuclear atoms return to their original positions, emitting energy as signals as they move back. The signals produce visible images for examination and diagnosis. MRI produces excellent contrast of soft tissues for evaluation of musculoskeletal conditions.
Analysis of synovial fluid may reveal inflammatory, septic, and noninflammatory joint diseases, which cause characteristic changes in the color, clarity, viscosity, and cellular elements of the fluid. The presence of blood in the joint fluid (hemarthrosis) usually indicates joint trauma. Normal synovial fluid is sterile, so the presence of bacteria in the fluid always indicates disease. Cell fragments and fibrous tissue in the fluid are the result of inflammation or wear and tear on the articular surfaces.
Tests of Muscular Function
When the individual’s history and physical examination disclose abnormalities, such as weakness, atrophy, muscle tenderness, cramps, and stiffness, specific tests of muscle function are in order. One of the most useful tests is the serum CK concentration. CK is found in large quantities in the muscle fibers, and when these are diseased or damaged, CK leaks into the serum. Myoglobin is also detectable in the urine after acute muscle damage caused by crush injury, ischemic disorders, extreme exertion, and some inherited diseases.
Because the muscle membrane tissue is excitable and carries an electrical charge, its capacity to function can be assessed by electromyography. Using sensitive needle electrodes, the electromyogram (EMG) records the summation of action potentials of the muscle fibers in each motor unit. The EMG is often compared with the electrocardiogram (ECG), but the activity recorded on the EMG is on a much smaller scale. The amplitude of the ECG is measured in volts, the duration of impulse is recorded in seconds, and both are recorded as the heart rate (e.g., 80 V/60 second). EMG amplitude is recorded in millivolts and the duration is measured in milliseconds, with a frequency of about 5 to 50 action potentials per second. Motor unit potentials are measured to determine rate of firing, duration, and amplitude. Abnormalities in EMG and nerve conduction velocities help differentiate muscle diseases (myopathy) from peripheral nerve (neuropathy) and neuromuscular junction disorders. The muscle biopsy (using histologic, histochemical, and electron microscopic studies) is used to further define the presence of myopathic and neuropathic disorders, many of which can be diagnosed only by muscle biopsy. Complex myography, a relatively new technique, allows a noninvasive way to gather information on the mechanical characteristics of muscle.20
A new area of evaluation is genetics. Recent advances in molecular genetics, deoxyribonucleic acid (DNA) libraries, genetic probes, and gene localization techniques have enhanced our knowledge of neuromuscular diseases, including types of muscular dystrophy, Charcot-Marie-Tooth disease, and familial amyotrophic lateral sclerosis.
Aging and the Musculoskeletal System
Aging is accompanied by the loss of bone tissue. Bones become less stiff, less strong, and more brittle with aging. The bone remodeling cycle takes longer to complete, and the rate of mineralization also slows down. Studies have shown that an individual’s bone mass at the end of their growth period helps determine the significance of bone loss as they age.12 With aging, women experience loss of bone density, accelerated by rapid bone loss during early menopause from increased osteoclastic bone resorption. By age 70, susceptible women have lost an average of 50% of their peripheral cortical bone mass. Extensive loss of bone mass leads to deformity, pain, stiffness, and a high risk for fractures. Men also experience bone loss but at later ages and much slower rates than women. Also, initial bone masses in men are approximately 30% higher than in women. However, the absolute risk for fracture is the same in men and women of the same age, and bone density is similar.12 Men’s peak bone mass is related to their race, heredity, hormonal factors (testosterone and estradiol), physical activity, and calcium intake during childhood. Bone loss in both sexes is related to smoking, calcium deficiency, magnesium deficiency, vitamin D deficiency, high protein intake, excess phosphorus intake, overly vigorous exercise, certain prescription and over-the-counter drug use, alcohol intake, and physical inactivity.21
Bone mass can be gained in healthy young women up to the third decade through physical activity, intake of dietary calcium, and magnesium. The positive effects of exercise in older adult women on bone mineral density (BMD) are not yet clear,22 but exercise has been shown to improve balance, coordination, muscle strength, lean body mass, and mobility, all of which may decrease the incidence of falls (also see Figure 2-22, p. 71). The use of oral contraceptives for maintaining bone mass is controversial. Height is also lost with aging because of increased spinal curvature, often because of asymptomatic vertebral fractures.
Aging of Joints
With aging, cartilage becomes more rigid, fragile, and susceptible to fibrillation because of more cross-linking of collagen and elastin, decreasing water content in the cartilage ground substance, and decreasing concentrations of glycosaminoglycans. Decreased range of motion of the joint is related to the changes in ligaments and muscles. Bones in joints develop evidence of osteoporosis with fewer trabeculae and thinner, less dense bones, making them prone to fractures. Intervertebral disk spaces decrease in height.
Aging of Muscles
The function of skeletal muscle depends on many factors that are affected by aging, including the nervous, vascular, and endocrine systems. In the young child the development of muscle tissue is highly dependent on continuing neurodevelopmental maturation. Muscle function remains trainable even into advanced age. Muscle diseases have a definite association with specific age groups. Muscular dystrophies occur in children, and muscle disabilities related to rheumatic diseases usually occur in advancing age.
Age-related loss in skeletal muscle is referred to as sarcopenia and is a direct cause of the age-related decrease in muscle strength. As the body ages, muscle bulk and strength decline slowly; thus strength is maintained into the 50s, with a slow decline in dynamic and isometric strength evident after age 70. Type II fibers decrease to a greater extent than the slower-acting type I fibers. Loss of satellite cells appears to play a major role in the development of sarcopenia.23 There is reduced RNA synthesis, loss of mitochondrial volume, and reduction in the size of motor units. The regenerative function of muscle tissue remains normal in aging persons. As much as 30% to 40% of skeletal muscle mass and strength may be lost from the third to ninth decades.
Maximal oxygen intake decreases with age. Reduced basal metabolic rate and decreased lean body mass are also seen in the older adult population.
α-Glycoprotein 1545
Actin 1559
Agonist 1562
Amphiarthrosis (slightly movable joint) 1549
Antagonist 1562
Appendicular skeleton 1546
Arthrography 1563
Arthroscopy 1563
Articular cartilage 1550
Axial skeleton 1546
Basement membrane 1558
Bone albumin 1545
Bone fluid 1545
Bone matrix 1541
Bone morphogenic protein (BMP) 1545
Bone-remodeling unit 1547
Calcification 1541
Canaliculi (sing, canaliculus) 1546
Chondrocyte 1551
Collagen fiber 1544
Compact bone (cortical bone) 1545
Contraction 1561
Coupling 1561
Creatine 1560
Creatine kinase 1560
Cross-bridge theory 1561
Diaphysis 1546
Diarthrosis (freely movable joint) 1549
Electromyogram (EMG) 1563
Endochondral ossification 1541
Endomysium 1555
Endosteum 1546
Epimysium 1555
Epiphyseal plate (growth plate) 1546
Epiphysis 1546
Excitation 1561
Fascia 1555
Fascicle 1555
Fibril 1544
Fibrous joint 1549
Flat bone 1547
Fusiform muscle 1555
Glycoprotein 1545
Glycosaminoglycan 1553
Golgi tendon organ 1557
Gomphosis 1549
Ground substance 1540
Growth plate 1546
Haversian canal 1546
Haversian system 1546
Hyaluronate 1550
Hydroxyapatite (HAP) 1545
Integrin 1544
Intima 1550
Intramembranous bone formation 1541
Irregular bone 1547
Isometric contraction (static or holding contraction) 1562
Isotonic contraction (lengthening or shortening) 1562
Joint (articulation) 1548
Joint capsule (articular capsule) 1550
Joint cavity (synovial cavity) 1550
Lacuna 1543
Lamellae 1546
Laminin 1545
Link protein 1553
Long bone 1546
Magnetic resonance imaging (MRI) 1563
Mesenchyme 1541
Metaphysis 1546
Motor unit 1556
Muscle cell 1557
Muscle fiber 1557
Muscle fiber action potential 1561
Muscle membrane 1558
Myoblast 1557
Myofibril 1557
Myosin 1559
Osteoblast 1542
Osteocalcin 1545
Osteoclast 1543
Osteocyte 1543
Osteoid 1542
Osteonectin 1545
Osteoprotegerin 1542
Oxygen debt 1562
Pennate muscle 1555
Perimysium 1555
Periosteum 1546
Procallus 1548
Protein core 1553
Proteoglycan 1545
RANKL (receptor activator of nuclear factor kB-ligand) 1543
Relaxation 1561
Remodeling 1547
Repetitive discharge 1562
Ruffled border 1544
Sarcolemma 1558
Sarcomere 1549
Sarcopenia 1564
Sarcoplasm 1558
Sarcoplasmic reticulum 1559
Sarcotubular system 1558
Sarcotubule 1559
Short bone (cuboidal bone) 1547
Sialoprotein (osteopontin) 1545
Skeletal (voluntary, striated, or extrafusal) muscle 1555
Sliding filament theory 1561
Spindle 1557
Spongy bone (cancellous bone) 1545
Subintima 1550
Suture 1549
Symphysis 1549
Synarthrosis (immovable joint) 1549
Synchondrosis 1549
Syndesmosis 1549
Synovial fluid 1550
Synovial joint (diarthrose) 1550
Synovial membrane (synovium) 1550
Tendon 1555
Tetanus 1562
Tidemark 1553
Trabeculae (sing., trabecula) 1546
Transverse tubule 1559
Type A synovial cell 1550
Type B synovial cell 1550
Type I fiber 1557
Type II fiber 1557
Woven bone (callus) 1548
REFERENCES
1. Hartmann, C. Skeletal development—Wnts are in control. Mol Cell. 2007;24(2):177.
2. Shapiro, F. Bone development and its relation to fracture repair. The role of mesenchymal osteoblasts and surface osteoblasts. Eur Cell Mater. 2008;15:53.
3. Glass, D.A., Karsenty, G. Minireview: in vivo analysis of Wnt signaling in bone. Endocrinol. 2007;148(6):2630.
4. Baron, R., Rawadi, G. Targeting the Wnt/beta-catenin pathway to regulate bone formation in the adult skeleton. Endocrinol. 2007;148(6):2635.
5. Issack, P.S., Helfet, D.L., Lane, J.M. Role of Wnt signaling in bone remodeling and repair. HSS J. 2008;4(1):66.
6. Caetano-Lopes, J., Canhão, H., Fonseca, J.E. Osteoblasts and bone formation. Acta Reum Port. 2007;32:103.
7. Yavropoulou, M.R., Yovos, J.G. The role of the Wnt signaling pathway in osteoblast commitment and differentiation. Hormones. 2008;6(4):279.
8. Zaudi, M., et al. Osteoclastogenesis, bone resorption, and osteoclast-based therapeutics. J Bone Mineral Res. 2003;18(4):599–609.
9. Boyce, B.F., Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res Ther. 2007;9(Suppl 1):S1.
10. Buckwalter J.A., Einhorn T.A., Simon S.R., eds. Orthopaedic basic science. Rosemont, IL: American Academy of Orthopaedic Surgeons, 1999.
11. Delaisse, J.M., et al. Matrix metalloproteinases (MMPs) and cathepsin K contribute differently to osteoclastic activities. Microsci Res Tech. 2003;61(6):504–513.
12. Seeman, E. Structural basis of growth-related gain and age-related loss of bone strength. Rheumatol. 2008;47:iv2.
13. Eyre, D.R., Weis, M.A., Wu, J.J. Articular cartilage collagen: an irreplaceable framework? Eur Cell Mater. 2006;12:57.
14. Jacobson, C., Duggan, D., Fishbach, G. Neuregulin induces the expression of transcription factors and myosin heavy chains typical of muscle spindles in cultured human muscle. Proc Natl Acad Sci U S A. 2004;101(33):12218–12223.
15. Labiet, D., et al. Calcium-dependent molecular spring elements in the giant protein titin. Proc Natl Acad Sci U S A. 2003;100(23):13716–13721.
16. Sargeant, A.J. Structural and functional determinants of human muscle power. Exp Physiol. 2007;92(2):323.
17. Richardson, R.S. Oxygen transport and utilization: an integration of the muscle systems. Adv Physiol Edu. 2003;27(1-4):183–191.
18. Abe, Y., Ishikawa, H., Fukao, A. Higher efficacy of urinary bone resorption marker measurements in assessing response to treatment for osteoporosis in postmenopausal women. Tohoku J Exp Med. 2008;214(1):51.
19. Ulrich-Vinther, M., et al. Articular cartilage biology. J Am Acad Orthoped Surg. 2003;11(6):421–430.
20. Rahe-Meyer, N., et al. Complex myograph allows the examination of complex muscle contractions for the assessment of muscle force, shortening, velocity, and work in vivo. Biomed Eng Online. 2008;7:20.
21. Karlsson, M. Does exercise reduce the burden of fractures? Acta Orthop Scand. 2002;73(6):691–705.
22. Moayyeri, A. The association between physical activity and osteoporotic fracture: a review of the evidence and implications for future research. Ann Epidemiol. 2008;18(11):827–835.
23. Verdijk, L.B., et al. Satellite cell content is specifically reduced in type II skeletal muscle fibers in the elderly. Am J Physiol Endocrinol Metab. 2007;292(1):E151.