Membranes and Transport
Learning objectives
After reading this chapter you should be able to:
 Describe differences between passive and active carrier-mediated transport systems.
Describe differences between passive and active carrier-mediated transport systems.
Describe the basic features of membrane channels and pores.
Give several specific examples of ion and substrate transport systems, including coupled transport systems.
Describe several characteristic diseases resulting from defects in membrane transport.
Introduction
Biomembranes are not rigid or impermeable, but highly mobile and dynamic structures
The plasma membrane is the gatekeeper of the cell. It controls not only the access of inorganic ions, vitamins and nutrients but also the entry of drugs and the exit of waste products. Integral transmembrane proteins have important roles in transporting these molecules through the membrane and often maintain concentration gradients across the membranes. K+, Na+ and Ca2+ concentrations in the cytoplasm are maintained at ∼140, 10, and 10−4 mmol/L, respectively, by transport proteins, whereas those outside (in the blood) are ∼5, 145, and 1–2 mmol/L, respectively. The driving force for transport of ions and maintenance of ion gradients is directly or indirectly provided by ATP. The transport properties of membranes will be illustrated by several important examples.
Types of transport processes
Simple diffusion through the phospholipid bilayer
Some small, neutral molecules can traverse biomembranes by simple diffusion
Small, nonpolar molecules (such as O2, CO2, N2) and uncharged polar molecules (such as urea, ethanol, and small organic acids) move through membranes by simple diffusion without the aid of membrane proteins (Table 8.1 and Fig. 8.1A). The direction of net movement of these species is always ‘downhill’, along the concentration gradient, from high to low concentration to establish equilibrium.
Table 8.1
Transport systems of biomembranes
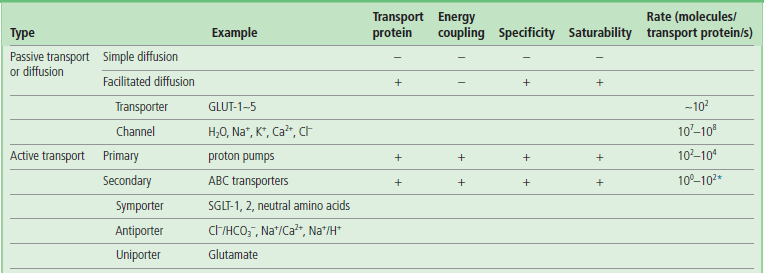
Transport systems are classified according to the role of transport proteins and energy coupling. Typical substrates for various types of transporters and channels are shown in the parentheses.
*The Cl−/HCO3− antiporter seems to be an exception to secondary active transport systems, as its transport rate is high, at 105 molecules/transport protein/s.
The hydrophobicity of the molecules is an important requirement for simple diffusion across the membrane, as the interior of the phospholipid bilayer is hydrophobic. The rate of transport of these molecules is, in fact, closely related to their partition coefficient between oil and water.
Although water molecules can be transported by simple diffusion, channel proteins (see below) are believed to control the movement of water across most membranes, especially in the kidney for concentration of the urine. Mutation in a water channel protein gene (aquaporin-2) causes diuresis in patients with nephrogenic diabetes insipidus, a disease characterized by excessive urination but without the hyper-glycemia characteristic of diabetes mellitus (see Chapter 23).
Transport mediated by membrane proteins
Membrane proteins are required for transport of larger molecules across biomembranes
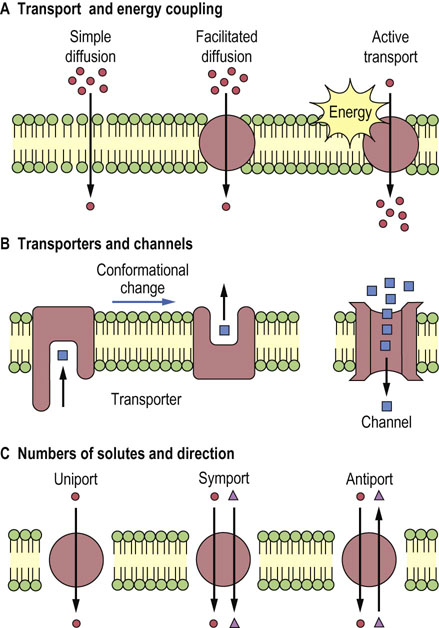
Transport of larger, polar molecules, such as amino acids or sugars, into a cell requires the involvement of membrane proteins known as transporters, also called porters, permeases, translocases or carrier proteins. The term ‘carrier’ is also applied to ionophores, which move passively across the membrane together with the bound ion (Fig. 8.2). Transporters are as specific as enzymes for their substrates, and work by one of two mechanisms: facilitated diffusion or active transport. Facilitated diffusion catalyzes the movement of a substrate through a membrane down a concentration gradient and does not require energy. In contrast, active transport is a process in which substrates are transported uphill, against their concentration gradient. Active transport must be coupled to an energy-producing reaction (see Fig. 8.1A).
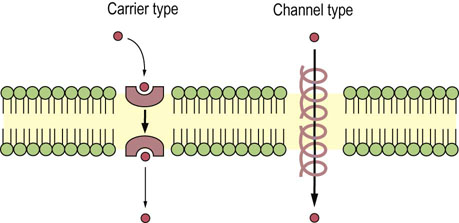
Fig. 8.2 Mobile ion carriers and channel-forming ionophores.
Ionophores permit net movement of ions only down their electrochemical gradients.
 Advanced concept box Antiobiotics and Membrane permeability
Advanced concept box Antiobiotics and Membrane permeability
Antibiotics act as ionophores and increase the permeability of membranes to specific ions; bactericidal effects of ionophores are attributed to disturbance of the ion transport systems of bacterial membranes. Ionophores permit net movement of ions only down their electrochemical gradients. There are two classes of ionophores: mobile ion carriers (or caged carriers) and channel formers (Fig. 8.2).
Valinomycin is a typical example of a mobile ion carrier. It is a cyclic peptide with a lipophilic exterior and an ionic interior. It dissolves in the membrane and diffuses between the inner and outer surfaces. K+ binds in the central core of valinomycin, and the complex diffuses across the membrane, releasing the K+, gradually dissipating the K+ gradient. The carrier-type ionophores, nigericin and monensin, exchange H+ for Na+ and K+, respectively. Ionomycin and A23187 are Ca2+ ionophores.
The β-helical gramicidin A molecule, a linear peptide with 15 amino acid residues, forms a pore. The head-to-head dimer of gramicidin A makes a transmembrane channel that allows movement of monovalent cations (H+, Na+, and K+).
Polyene antibiotics such as amphotericin B and nystatin exert their cytotoxic action by rendering the membrane of the target cell permeable to ions and small molecules. Formation of a sterol–polyene complex is essential for the cytotoxic function of these antibiotics, as they display a selective action against organisms in which the membranes contain sterols. Thus they are active against yeasts, a wide variety of fungi, and other eukaryotic cells, but have no effect on bacteria. Because their affinity toward ergosterol, a fungal membrane component, is higher than that for cholesterol, these antibiotics have been used for the treatment of topical infections of fungal origin.
Saturability and specificity are important characteristics of membrane transport systems
The rate of facilitated diffusion is generally much greater than that of simple diffusion: transport proteins catalyze the transport process. In contrast to simple diffusion, in which the rate of transport is directly proportional to the substrate concentration, facilitated diffusion is a saturable process, having a maximum transport rate, Tmax (Fig. 8.3). When the concentration of extracellular molecules (transport substrates) becomes very high, the Tmax is achieved by saturation of the transport proteins with substrate. The kinetics of facilitated diffusion for substrates can be described by the same equations that are used for enzyme catalysis (e.g. Michaelis–Menten and Lineweaver–Burk type equations) (see Chapter 6):

Fig. 8.3 Comparison of the transport kinetics of facilitated diffusion and simple diffusion.
The rate of transport of substrate is plotted against the concentration of substrate in the extracellular medium. In common with enzyme catalysis, transporter-catalyzed uptake has a maximum transport rate, Tmax (saturable). Kt is the concentration at which the rate of substrate uptake is half-maximal. For simple diffusion, the transport rate is slower and directly proportional to substrate concentration.
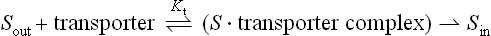
where Kt is the dissociation constant of the substrate–transporter complex, and Sout is the concentration of transport substrate. Then the transport rate, t, can be calculated as:
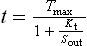
where the Kt is the concentration that gives the half-maximal transport rate. The Kt for a transporter is conceptually the same as the Km for an enzyme (Chapter 6).
The transport process is usually highly specific: each transporter transports only a single species of molecules or structurally related compounds. The red blood cell GLUT-1 transporter has a high affinity for D-glucose, but 10–20 times lower affinity for the related sugars, D-mannose and D-galactose. The enantiomer L-glucose is not transported; its Kt is more than 1000 times higher than that of the D-form.
 Clinical box Cystinosis
Clinical box Cystinosis
An 18-month-old child presented with polyuria, failure to thrive and an episode of severe dehydration. Urine dipstick testing demonstrated glucosuria and proteinuria, with other biochemical analyses showing generalized aminoaciduria and phosphaturia.
This is a classical presentation of infantile cystinosis, resulting from accumulation of cystine in lysosomes because of a defect in the lysosomal transport protein, cystinosine. Cystine is poorly soluble, and crystalline precipitates form in cells throughout the body. In vitro experiments with cystine loading have shown that renal proximal tubular cells become ATP-depleted, resulting in impairment of ATP-dependent ion pumps with consequent electrolyte imbalances and metabolite losses.
Treatment with cysteamine increases the transport of cystine from lysosomes, delaying the decline in renal function. Cysteamine is a weak base; it forms a mixed disulfide with cysteine, which is then secreted through a cationic amino acid transporter. If untreated, renal failure occurs by 6–12 years of age. Unfortunately, there is further accumulation of cystine in the central nervous system, despite therapy, with long-term neurological damage.
Clinical box Hartnup disease
A three-year old had been on holiday to Spain and developed pellagra-like skin changes on his face, neck, forearms and dorsal aspects of his hands and legs. His skin became scaly, rough and hyperpigmented. The child was brought to the GP complaining of headaches and weakness. Urinalysis demonstrated gross hyperaminoaciduria of neutral monoamino-monocarboxylic acids (i.e. alanine, serine, threonine, asparagine, glutamine, valine, leucine, isoleucine, phenylalanine, tyrosine, tryptophan, histidine and citrulline).
These amino acids share a common transporter which is expressed only on the luminal border of epithelial cells in the renal tubules and intestinal epithelium. The pellagra-like dermatitis (see Chapter 11) and neurological involvement resemble nutritional niacin deficiency. The reduced tryptophan intake results in reduced nicotinamide production. The disease is treated with oral nicotinamide and application of sun-blocking agents to exposed areas.
Characteristics of glucose transporters (uniporters)
Glucose transporters catalyze downhill transport of glucose into and out of cells
Glucose transporters are essential for facilitated diffusion of glucose into cells. The GLUT family of glucose transporters includes GLUT-1 to GLUT-5 (Table 8.2) and others. They are transmembrane proteins similar in size, all having about 500 amino acid residues and 12 transmembrane helices. GLUT-1, in red blood cells, has a Km of ∼2 mmol/L. The GLUT-1 transporter operates at about 40% of Tmax under fasting conditions (blood glucose concentration of 5 mmol/L; 90 mg/dL); this level of activity is sufficient to meet the needs of the red cell (Chapter 12). In contrast, pancreatic islet β-cells express GLUT-2, with a Km of more than 10 mmol/L (180 mg/dL). In response to the intake of food and the resulting increase in blood glucose concentration, GLUT-2 molecules respond by increasing the uptake of glucose into β-cells, stimulating insulin secretion (Chapter 21). Cells in insulin-sensitive tissues such as muscle and adipose have GLUT-4. Insulin stimulates translocation of GLUT-4 from intracellular vesicles to the plasma membrane, facilitating glucose uptake during meals.
Table 8.2
Classification of glucose transporters
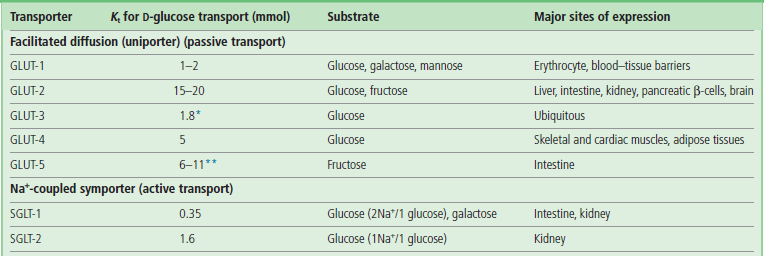
Km values are determined from the uptake of 2-deoxy-D-glucose (*), a nonmetabolizable analogue of glucose, and fructose (**).
Clinical box Defective glucose transport across the blood–brain barrier as a cause of seizures, and developmental delay
A male infant at the age of 3 months suffered from recurrent seizures. His cerebrospinal fluid (CSF) glucose concentrations were low (0.9–1.9 mmol/L; 16–34 mg/dL), and the ratio of CSF to blood glucose ranged from 0.19 to 0.33; the normal value is 0.65.
The potential causes of low CSF glucose concentrations, such as bacterial meningitis, subarachnoid hemorrhage, and hypoglycemia, were not present, and high CSF lactate values would be found in all these conditions except hypoglycemia. In contrast, the CSF lactate concentrations were consistently low (0.3–0.4 mmol/L; 3–4 mg/dL) compared with the normal value (<2.2 mmol/L; <20 mg/dL). These findings suggested a defect in transport of glucose from the blood to the brain.
Assuming that the activity of GLUT-1 glucose transporter in the erythrocyte reflects that of the brain microvessels, a transport assay using his erythrocytes was carried out. The Tmax for uptake of glucose by the patient's erythrocytes was 60% of the mean normal value, suggesting a heterozygous defect. A ketogenic diet (a high-fat, low-protein, low-carbohydrate diet) was started, since the brain can use ketone bodies as oxidizable fuel sources, and the entry of ketone bodies into the brain is not dependent on the glucose transporter system. The patient stopped having seizures within 4 days after beginning the diet.
Transport by channels and pores
Membrane channels or pores are open, less selective conduits for transport of ions, metabolites and even proteins across biomembranes
Channels are often pictured as tunnels across the membrane, in which binding sites for substrates (ions) are accessible from either side of the membrane at the same time (see Fig. 8.1B). Conformational changes are not required for the translocation of substrates entering from one side of the membrane to exit on the other side. However, voltage changes and ligand binding induce conformational changes in channel structure that have the effect of opening or closing the channels – processes known as voltage or ligand ‘gating’. Movement of molecules through channels is fast in comparison with the rates achieved by transporters (see Table 8.1).
The terms ‘channel’ and ‘pore’ are sometimes used interchangeably. However, ‘pore’ is used most frequently to describe more open, somewhat nonselective structures that discriminate between substrates, e.g. peptides or proteins, on the basis of size. The term ‘channel’ is usually applied to more specific ion channels.
Three examples of pores important for cellular physiology
The gap junction between endothelial, muscle, and neuronal cells is a cluster of small pores, in which two cylinders of six connexin subunits in the plasma membranes join each other to form a pore about 1.2–2.0 nm (12–20 Å) in diameter. Molecules smaller than about 1 kDa can pass between cells through these gap junctions. Such cell–cell interchange is important for physiologic communication or coupling, for example in the concerted contraction of uterine muscle during labor and delivery. Mutations of the genes encoding connexin 26 and connexin 32 cause deafness and Charcot–Marie–Tooth disease, respectively.
Nuclear pores have a radius of about 9.0 nm (90 Å) through which larger proteins and nucleic acids enter and leave the nucleus.
A third class of pores is important for protein sorting. Mitochondrial proteins encoded by nuclear genes are transported to this organelle through pores in the outer mitochondrial membrane. Nascent polypeptide chains of secretory proteins and plasma membrane proteins also pass through pores in the endoplasmic reticulum membrane during biosynthesis of the peptide chain.
Active transport
Primary active transport systems use ATP directly to drive transport; secondary active transport uses an electrochemical gradient of Na+ or H+ ions, or a membrane potential produced by primary active transport processes
ATP is a high-energy product of metabolism and is often described as the ‘energy currency’ of the cell (Chapter 9). The phosphoanhydride bond of ATP releases free energy when it is hydrolyzed to produce adenosine diphosphate (ADP) and inorganic phosphate. Such energy is used for biosynthesis, cell movement, and uphill transport of molecules against concentration gradients. Primary active transport systems use ATP directly to drive transport; secondary active transport uses an electrochemical gradient of Na+ or H+ ions, or a membrane potential produced by primary active transport processes. Sugars and amino acids are generally transported into cells by secondary active transport systems.
Primary active transport systems use ATP to drive ion pumps (ion transporting ATPases or pump ATPases)
The pump ATPases are classified into four groups (Table 8.3). Coupling factor ATPases (F-ATPases) in mitochondrial, chloroplast, and bacterial membranes hydrolyze ATP and transport hydrogen ions (H+). As discussed in detail in the next chapter, the mitochondrial F-ATPase works in the backward direction, synthesizing ATP from ADP and phosphate as protons move down an electrochemical (concentration and charge) gradient generated across the inner mitochondrial membrane during oxidative metabolism. The product, ATP, is released into the mitochondrial matrix, but is needed for biosynthetic reactions in the cytoplasm. ATP is transported to the cytoplasm through an ATP-ADP translocase in the mitochondrial inner membrane. This translocase is an example of an antiport system (see Fig. 8.1C); it allows one molecule of ADP to enter only if one molecule of ATP exits simultaneously.
Table 8.3
Primary active transporters in eukaryotic cells
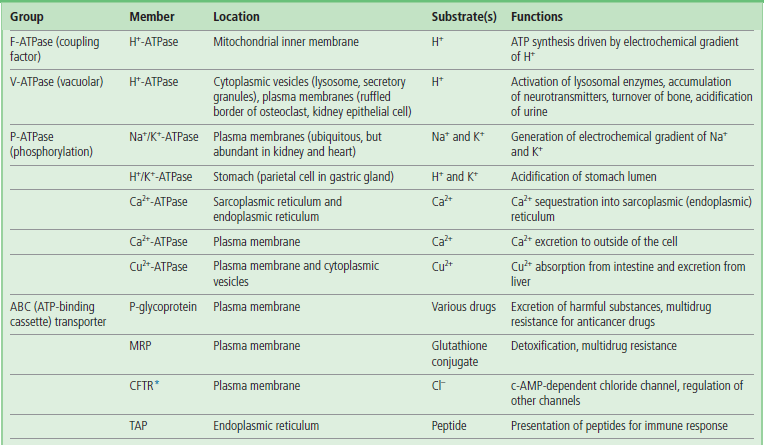
Various examples of primary active transporters (ATP-powered pump ATPases) are listed, together with their location.
*Some ABC transporters function as channels or channel regulators. MRP, multidrug resistance-associated protein; CFTR, cystic fibrosis transmembrane conductance regulator; TAP, transporter associated with antigen processing.
Cytoplasmic vesicles, such as lysosomes, endosomes, and secretory granules, are acidified by a V-type (vacuolar) H+-ATPase in their membranes. Acidification by this V-ATPase is important for the activity of lysosomal enzymes that have acidic pH optima, and for the accumulation of drugs and neurotransmitters in secretory granules. The V-ATPase also acidifies the extracellular environments of osteoclasts and renal epithelial cells. Defects in the osteoclast plasma membrane V-ATPase result in osteopetrosis (increased bone density), while mutation of the ATPase in collecting ducts of the kidney causes renal tubular acidosis. F- and V-type ATPases are structurally similar, and seem to be derived from a common ancestor. The ATP-binding catalytic subunit and the subunit forming the H+ pathway are conserved between these ATPases.
P-ATPases form phosphorylated intermediates that drive ion translocation: the ‘P’ refers to the phosphorylation. These transporters have an active-site aspartate residue that is reversibly phosphorylated by ATP during the transport process. The P-type Na+/K+-ATPase in various tissues and the Ca2+-ATPase in the sarcoplasmic reticulum have important roles in maintaining cellular ion gradients. Na+/K+-ATPases also create an electrochemical gradient of Na+ that produces the driving force for uptake of nutrients from the intestine (below). The discharge of this electrochemical gradient is also fundamental to the process of nerve transmission. Mutations of P-ATPase genes cause Brody cardiomyopathy (Ca2+-ATPase), familial hemiplegic migraine type 2 (Na+/K+-ATPase), and Menkes and Wilson's diseases (Cu2+-ATPases).
Clinical box Menkes and wilson's diseases
X-linked Menkes disease is a lethal disorder that occurs in 1 in 100,000 newborn infants and is characterized by abnormal and hypopigmented hair, a characteristic facies, cerebral degeneration, connective tissue and vascular defects, and death by the age of 3 years. A copper-transporting P-ATPase that is expressed in all tissues except liver is defective in this disease (Table 8.3). In patients with Menkes disease, copper enters the intestinal cells, but is not transported further, resulting in severe copper deficiency. Subcutaneous administration of a copper histidine complex may be an effective treatment if started early.
The gene for Wilson's disease also encodes a copper-transporting P-ATPase and is 60% identical with that of the Menkes gene. It is expressed in liver, kidney, and placenta. Wilson's disease occurs in 1 in 35,000–100,000 newborns and is characterized by failure to incorporate copper into ceruloplasmin in the liver and failure to excrete copper from the liver into bile, resulting in toxic accumulation of copper in the liver and also in the kidney, brain, and cornea. Liver cirrhosis, progressive neurologic damage, or both, occur during childhood to early adulthood. Chelating agents such as penicillamine are used for treatment of patients with this disease. Oral zinc treatment may be useful for decreasing the absorption of dietary copper. Copper is an essential trace metal and an integral component of many enzymes. However, it is toxic in excess, because it binds to proteins and nucleic acids, enhances the generation of free radicals, and catalyzes oxidation of lipids and proteins in membranes.
The ATP-binding cassette (ABC) transporters comprise the fourth active transporter family. ‘ABC’ is the abbreviation for ‘ATP-binding cassette’, referring to an ATP-binding region in the transporter (see Table 8.3). P-glycoprotein (‘P’ = permeability) and MRP (multidrug resistance-associated protein), which have a physiologic role in excretion of toxic metabolites and xenobiotics, contribute to resistance of cancer cells to chemotherapy. TAP transporters, a class of ABC transporters associated with Antigen Presentation, are required for initiating the immune response against foreign proteins; they mediate antigen peptide transport from the cytosol into endoplasmic reticulum. Some ABC transporters are present in peroxisomal membrane where they appear to be involved in the transport of peroxisomal enzymes necessary for oxidation of very long-chain fatty acids. Defects of ABC transporters are associated with a number of diseases (see box on next page).
Advanced concept box ABC transporter diseases
Human genome data suggests that there are about 50 genes for ABC transporters. An unusually wide range of diseases are caused by defects in ABC transporters, including Tangier disease, Stargardt disease, progressive intrahepatic cholestasis, Dubin–Johnson syndrome, pseudoxanthoma elasticum, familial persistent hyperinsulinemic hypoglycemia of infancy (PHHI), adrenoleukodystrophy, Zellweger syndrome, sitosterolemia and cystic fibrosis.
Cystic fibrosis (CF) is the most common potentially lethal autosomal recessive disease of Caucasian populations, affecting 1 in 2500 newborns. CF is usually manifested as exocrine pancreatic insufficiency, an increase in the concentration of chloride ions (Cl–) in sweat, male infertility, and airway disease, which is the major cause of morbidity and mortality. The pancreatic and lung pathology results from the increased viscosity of secreted fluids (mucoviscoidosis). CF is caused by mutations in the gene CFTR (cystic fibrosis transmembrane conductance regulator) which encodes a Cl– channel. ATP binding to CFTR is required for channel opening. The lack of this channel activity in epithelia of CF patients affects both ion and water secretion.
Uniport, symport, and antiport are examples of secondary active transport
Transport processes may be classified into three general types: uniport (monoport), symport (cotransport) and antiport (countertransport) (see Fig. 8.1). Transport substrates move in the same direction during symport, and in opposite directions during antiport. Uniport of charged substrates may be electrophoretically driven by the membrane potential of the cell. The movement of one substrate uphill, against its concentration gradient, can be driven by antiport of another substrate (usually a cation such as Na+ or H+) down a gradient. The proteins participating in these transport systems are termed uniporters, symporters, and antiporters, respectively (see Table 8.1). Some examples are presented below.
Advanced concept box Electrochemical gradients
The permeability of most nonelectrolytes through membranes can be analyzed by assuming that the rate-limiting step is the diffusion within the lipid bilayer. Their permeability across a phospholipid bilayer is experimentally shown to be a function of the partition coefficient between aqueous and organic solvents. The relative rate of simple diffusion of a molecule across the membrane is therefore proportional to the concentration gradient across the bilayer and to the hydrophobicity of the molecule.
For charged molecules and ions, transport across the membrane must be facilitated by a transporter or channel, and is driven by the electrochemical gradient, a combination of the concentration gradient (chemical potential) and the voltage gradient across the membrane (electric potential). These forces may act in the same direction or in opposite directions.
In the case of Na+ ions, the concentration difference between outside (145 mmol/L) and inside (12 mmol/L) the cell is about a factor of 10, being maintained by the Na+/K+-ATPase. The Na+/K+-ATPase is electrogenic, pumping out three Na+ and pumping in two K+ ions, generating an inside-negative membrane potential. K+ leaks out through K+ channels, down its concentration gradient (140 mmol/L to 5 mmol/L), further increasing the electric potential. The concentration gradient of Na+ ions and the electric potential (inside negative) power the import and export of other molecules with Na+ against their concentration gradient by symporters and antiporters, respectively.
Examples of transport systems and their coupling
Ca2+ transport and mobilization in muscle
Membrane depolarization opens up voltage dependent ion channels at the neuromuscular junction
Striated muscle (skeletal and cardiac) is composed of bundles of muscle cells (Chapter 20). Each cell is packed with bundles of actin and myosin filaments (myofibrils) that produce contraction. During muscle contraction, nerves at the neuromuscular junction stimulate local depolarization of the membrane by opening voltage-dependent Na+ channels. The depolarization spreads rapidly into invaginations of the plasma membrane called the transverse (T) tubules, which extend around the myofibrils (see Fig. 20.5).
Voltage-dependent Ca2+ channels (VDCCs) located in the T tubules of skeletal muscle change their conformation in response to membrane depolarization, and directly activate a Ca2+-release channel in the sarcoplasmic reticulum membrane, a network of flattened tubules that surrounds each myofibril in the muscle cell cytoplasm. The escape of Ca2+ from the lumen (interior compartment) of the sarcoplasmic reticulum increases the cytoplasmic concentration of Ca2+ (depolarization-induced Ca2+ release) about 100-fold, from 10−4 mmol/L (0.0007 mg/L) to about 10−2 mmol/L (0.07 mg/dL), triggering ATP hydrolysis by myosin, which initiates muscle contraction. A Ca2+-ATPase in the sarcoplasmic reticulum then hydrolyzes ATP to transport Ca2+ back out of the cytoplasm into the lumen of the sarcoplasmic reticulum, decreasing the cytoplasmic Ca2+ and allowing the muscle to relax (Fig. 8.4, left).
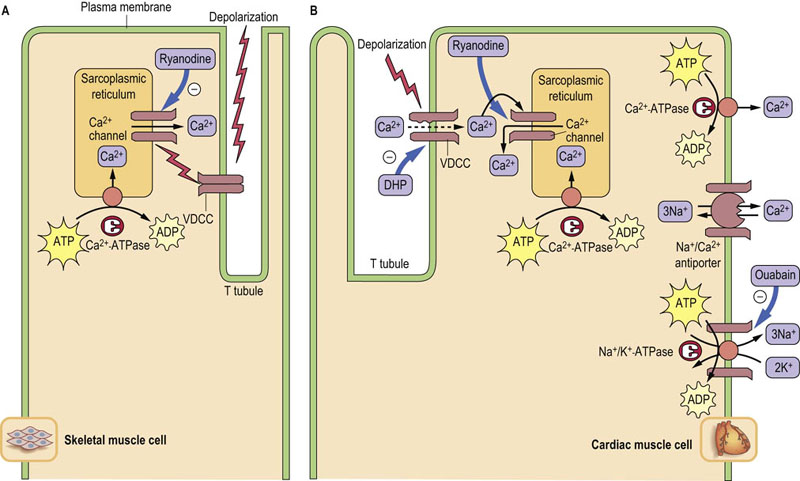
Fig. 8.4 Ca2+ movement in muscle contraction cycle.
Roles of transporters in Ca2+ movements in skeletal (A) and cardiac (B) muscle cells during contraction. Thick arrows indicate the binding sites for inhibitors. In skeletal muscle, VDCCs directly activate release of Ca2+ from the sarcoplasmic reticulum. The increased cytoplasmic Ca2+ concentration triggers muscle contraction. A Ca2+-ATPase in the sarcoplasmic reticulum pumps Ca2+ back into the lumen, decreasing the cytoplasmic Ca2+ concentration, and the muscle relaxes. In heart muscle, VDCCs allow entry of a small amount of Ca2+, which induces release of Ca2+ from the lumen of sarcoplasmic reticulum. Two types of Ca2+-ATPases and an Na+/Ca2+-antiporter are responsible for pumping cytoplasmic Ca2+ out of the muscle cell. The Na+/Ca2+-antiporter uses the sodium (Na+) gradient produced by Na+/K+-ATPase to antiport Ca2+. DHP, dihydropyridine, nifedipine, a calcium channel blocker used for treatment of hypertension. Ryanodine is a potent inhibitor of the Ca++ channel in the sarcoplasmic reticulum.
In cardiac muscle, VDCCs permit the entry of a small amount of Ca2+, which then stimulates Ca2+ release through the Ca2+ channel from the lumen of the sarcoplasmic reticulum (Ca2+-induced Ca2+ release). Not only the sarcoplasmic reticulum Ca2+-ATPase but also an Na+/Ca2+-antiporter and a plasma membrane Ca2+-ATPase are responsible for pumping out cytoplasmic Ca2+ from heart muscle (Fig. 8.4, right). The rapid restoration of ion gradients allows for rhythmic contraction of the heart.
Active transport of glucose in the intestines
A Na+/K+-ATPase drives uptake of glucose into intestinal and renal epithelial cells
The transport of blood glucose into cells is generally by facilitated diffusion, as the intracellular concentration of glucose is typically less than that of blood (see Table 8.2). In contrast, the transport of glucose from the intestine into blood involves both facilitated diffusion and active transport processes (Fig. 8.5). Active transport is especially important for maximal recovery of sugars from the intestine when the intestinal concentration of glucose falls below that in the blood.
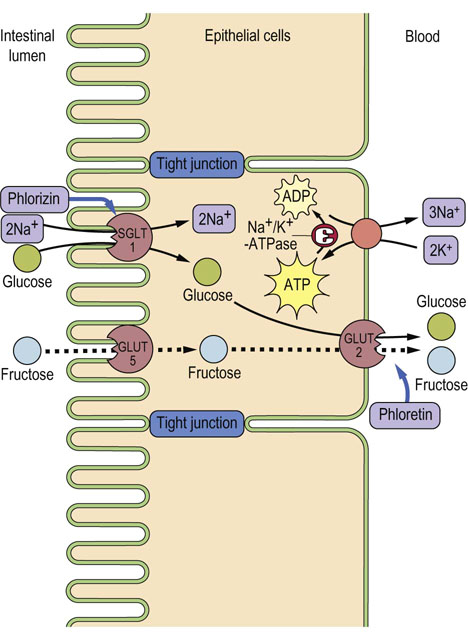
Fig. 8.5 Glucose transport from intestinal lumen into the blood.
Glucose is pumped into the cell through the Na+-coupled glucose symporter (SGLT1), and passes out of the cell by facilitated diffusion mediated by the GLUT-2 uniporter. The Na+ gradient for glucose symport is maintained by the Na+/K+-ATPase, which keeps the intracellular concentration of Na+ low. SGLT1 is inhibited by phlorizin and GLUT-2 by phloretin. Phloretin-insensitive GLUT-5 catalyzes the uptake of fructose by facilitated diffusion. The fructose is then exported through GLUT-2. A defect of SGLT1 causes glucose/galactose malabsorption. Adjacent cells are connected by impermeable tight junctions, which prevent solutes from crossing the epithelium.
An Na+-coupled glucose symporter SGLT1, driven by a Na+ gradient formed by Na+/K+-ATPase, transports glucose into the intestinal epithelial cell, while GLUT-2 facilitates the downhill movement of glucose into the portal circulation (see Fig. 8.5). A similar pathway operates in the kidney.
The renal glomerulus is an ultrafiltration system that filters small molecules from blood. However, glucose, amino acids, many ions, and other nutrients in the ultrafiltrate are almost completely reabsorbed in the proximal tubules, by symport processes. Glucose is reabsorbed primarily by sodium glucose transporter 2 (SGLT2; one-to-one Na+:Glc stoichiometry) into renal proximal tubular epithelial cells. Much smaller amounts of glucose are recovered by SGLT1 in a later segment of the tubule, which couples transport of one molecule of glucose to two sodium ions. The concentration of Na+ in the filtrate is 140 mmol/L (322 mg/dL), while that inside the epithelial cells is 30 mmol/L (69 mg/dL), so that Na+ flows ‘downhill’ along its gradient, dragging glucose ‘uphill’ against its concentration gradient. As in intestinal epithelial cells, the low intracellular concentration of Na+ is maintained by an Na+/K+-ATPase on the opposite side of the tubular epithelial cell, which antiports three cytoplasmic sodium ions for two extracellular potassium ions, coupled with hydrolysis of a molecule of ATP.
Clinical box Modulation of transporter activity in diabetes
The ATP-sensitive K+ channel (KATP) participates in regulation of insulin secretion in pancreatic islet β-cells. When the blood concentration of glucose increases, glucose is transported into the β-cell through a glucose transporter (GLUT-2) and metabolized, resulting in an increase in cytoplasmic ATP concentration. The ATP binds to the regulatory subunit of the K+ channel, KATP-β (called the sulfonylurea receptor, SUR1) causing structural change of a KATP-α subunit, which closes the KATP channel. This induces depolarization of the plasma membrane (decreased voltage gradient across the membrane) and activates voltage-dependent calcium (Ca2+) channels (VDCCs). The entry of Ca2+ stimulates exocytosis of vesicles that contain insulin.
The binding of sulfonylureas such as tolbutamide and glibenclamide to KATP-β on the outside of the plasma membrane is thought to mimic the regulatory effect of intracellular ATP. Sulfonylureas stimulate insulin secretion, which decreases blood glucose concentration in diabetes.
Defective KATP channels, which are unable to transport K+, induce low blood glucose concentration – a condition called Persistent Hyperinsulinemic Hypoglycemia of Infancy (PHHI) that occurs in 1 per 50,000 persons – as a result of loss of K+-channel function and continuous insulin secretion.
Advanced concept box Various drugs inhibit transporters in muscle
Phenylalkylamine (verapamil), benzothiazepine (diltiazem), and dihydropyridine (DHP; nifedipine) are Ca2+-channel blockers that inhibit VDCCs (Fig. 8.4). Ryanodine inhibits the Ca2+-release channel in the sarcoplasmic reticulum. These drugs are used as antihypertensive agents to inhibit the increase in cytoplasmic Ca2+ concentration and thus the force of muscle contraction. In contrast, cardiac glycosides such as ouabain and digoxin increase heart muscle contraction and are used for treatment of congestive heart failure. They act by inhibiting the Na+/K+-ATPase that generates the Na+ concentration gradient used to drive export of Ca2+ by the Na+/Ca2+ antiporter. Snake venoms such as α-bungarotoxin, and tetrodotoxin from the puffer fish inhibit voltage-dependent Na+ channels. Lidocaine, a Na+-channel blocker, is used as a local anesthetic and antiarrhythmic drug. Inhibition of Na+ channels represses transmission of the depolarization signal.
Acidification of gastric juice by a proton pump in the stomach
A P-ATPase in gastric parietal cells maintains the low pH of the stomach
The lumen of the stomach is highly acidic (pH ≈1) because of the presence of a proton pump (H+/K+-ATPase; P-ATPase in Table 8.3) that is specifically expressed in gastric parietal cells. The gastric proton pump is localized in intracellular vesicles in the resting state. Stimuli such as histamine and gastrin induce fusion of the vesicles with the plasma membrane (Fig. 8.6A). The pump antiports two cytoplasmic protons and two extracellular potassium ions, coupled with hydrolysis of a molecule of ATP; thus it is called an H+/K+-ATPase. The counter-ion C1– is secreted through a Cl– channel, producing hydrochloric acid (HCl) (gastric acid) in the lumen (Fig. 8.6B).
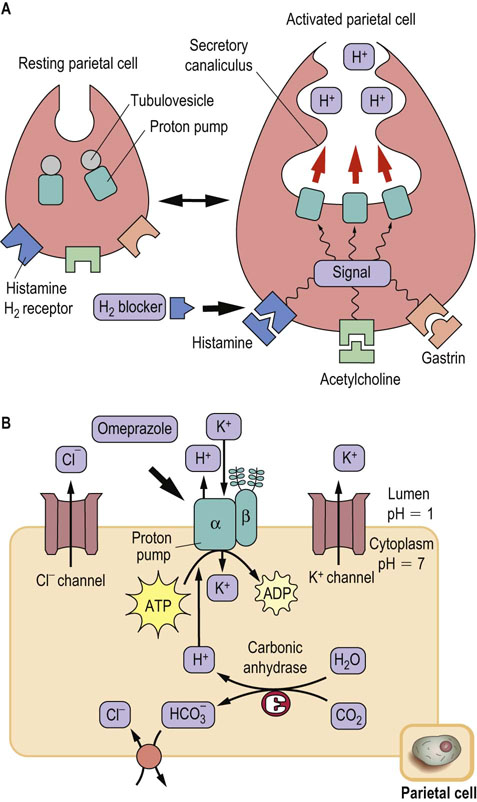
Fig. 8.6 Acid secretion from gastric parietal cells.
(A) Acid secretion is stimulated by extracellular signals and accompanied by morphologic changes in parietal cells, from resting (left) to activated (right). The proton pump (H+/K+-ATPase) moves to the secretory canaliculus (plasma membrane) from cytoplasmic tubulovesicles. H2-blockers compete with histamine at the histamine H2-receptor. (B) Ion balance in the parietal cell. The H+ transported by the proton pump are supplied by carbonic anhydrase. Bicarbonate, the other product of this enzyme, is antiported with Cl–, which is secreted through a Cl– channel. The potassium ions imported by the proton pump are again excreted by a K+ channel. The proton pump has catalytic α- and glycosylated β-subunits. The drug omeprazole covalently modifies cysteine residues located in the extracytoplasmic domain of the α-subunit and inhibits the proton pump. Thick arrows indicate the binding sites for inhibitors.
Renal Na+ transport and renin–angiotensin–aldosterone system
The kidney is a major site for regulation of Na+ homeostasis; several transport systems participate in Na+ handling in the kidney tubules
The kidney plays important parts in preservation of water and electrolyte homeostasis (Chapter 23). The primary urine is formed by glomerular filtration. Salts passing through the glomerular filter are reabsorbed in the renal tubule. Most of the Na+ reabsorption in the tubules occurs in proximal segments. Although <10% of Na+ reabsorption occurs in the distal nephron, this site is recognized to be essential for fine-tuning of plasma volume since Na+ reabsorption in the distal nephron is sensitive to the steroid hormone aldosterone.
As indicated in Figure 8.7, several Na+ transport proteins participate in Na+ reabsorption in the kidney tubules. Among them, Na+/K+-ATPase expressed in the basolateral membranes of epithelial cells transports Na+ across the membrane into the blood and generates a low intracellular Na+ concentration. The Na+ ions in the urine are transported into the epithelial cells through apical membranes by segment-specific Na+ transport proteins. Na+/H+-antiporter (NHE3) mediates Na+ entry in the proximal tubule. Furosemide-sensitive Na+-K+-2Cl–-symporter (NKCC2) and thiazide-sensitive Na+-Cl–-cotransporter (NCCT) are localized in thick ascending loop of Henle and distal convoluted tubules, respectively. Amiloride-sensitive epithelial Na+ channel (ENaC) is expressed in the aldosterone-sensitive distal nephron. The ionic balance of epithelial cells is also regulated by the renal outer medullary K+ channel (ROMK) and the Cl– channel (ClC).
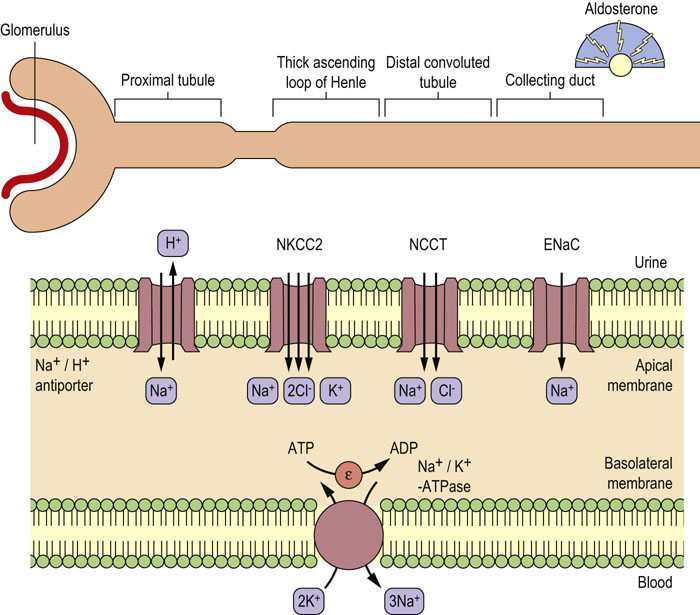
Fig. 8.7 Segment-specific expression of renal Na+ transport proteins.
Various Na+ transport proteins expressed in the apical membranes of renal epithelial cells transport Na+ from the urine into the cells. Na+ in the cells is excreted into the blood by an Na+/K+-ATPase expressed in the basolateral membranes. K+ ions are recycled through basolateral ROMK in the thick ascending loop of Henle and aldosterone sensitive distal nephron. Chloride ions leaves the cells in the thick ascending loop of Henle and distal convoluted tubule through ClCs.
The renin-angiotensin-aldosterone system plays a central role in the regulation of circulating volume, extracellular fluid osmolality and blood pressure
As discussed in more detail in Chapters 24 and 25, renin is produced in the juxtaglomerular apparatus of the kidney upon decrease of circulating volume, and it converts angiotensinogen produced in the liver to angiotensin I. Angiotensin I is converted to angiotensin II by angiotensin-converting enzyme (ACE). Angiotensin II produced in the blood promotes the release of aldosterone from the adrenal glands. Aldosterone promotes renal Na+ and water retention and elevated blood pressure by increasing Na+ reabsorption in the cortical collecting duct. Angiotensin II also promotes elevations in blood pressure by direct vasoconstriction and by increasing Na+ reabsorption in the proximal tubule.
Clinical box Inhibiting the gastric proton pump and eradication of helicobacter pylori
Chronic strong acid secretion by the gastric proton pump injures the stomach and the duodenum, leading to gastric and duodenal ulcers. Proton pump inhibitors such as omeprazole are delivered to parietal cells from the circulation after oral administration. Omeprazole is a prodrug: it accumulates in the acidic compartment, as it is a weak base, and is converted to the active compound under the acidic conditions in the gastric lumen. The active form covalently modifies cysteine residues located in the extracytoplasmic domain of the proton pump. H2-blockers (receptor antagonists) such as cimetidine and ranitidine indirectly inhibit acid secretion by competing with histamine for its receptor (Fig. 8.6).
Infection of the stomach by Helicobacter pylori also causes ulcers and is associated with an increased risk of gastric adenocarcinoma. Recently, antibiotic treatment has been introduced to eradicate H. pylori. Interestingly, antibiotic treatment together with omeprazole is much more effective, possibly because of an increased stability of the antibiotic under the weakly acidic condition produced by proton pump inhibition.
Summary
Most of the permeability properties of the membrane are determined by transport proteins, which are integral membrane proteins.
Protein-mediated transport is a saturable process with high substrate specificity.
Facilitated diffusion is catalyzed by transporters that permit the movement of ions and molecules down concentration gradients, whereas uphill or active transport requires energy.
Primary active transport is catalyzed by pump ATPases that use energy produced by ATP hydrolysis.
Secondary active transport uses electrochemical gradients of Na+ and H+, or membrane potential produced by primary active transport processes. Uniport, symport, and antiport are examples of secondary active transport.
Numerous substrates such as ions, nutrients, small organic molecules including drugs and peptides, and proteins are transported by various transporters.
All these transporters are indispensable for homeostasis. The expression of unique sets of transporters is important for specific cell functions such as muscle contraction, nutrient and ion absorption by intestinal epithelial cells, resorption of nutrients by kidney cells, and secretion of acid from gastric parietal cells.
Camargo, SM, Brockenhauer, D, Kleta, R. Aminoacidurias: clinical and molecular aspects. Kidney Int. 2008; 73:918–925.
Hinz, A, Tampe, R. ABC transporters and immunity: mechanism of self-defense. Biochemistry. 2012; 51:4981–4989.
Linton, KJ. Structure and function of ABC transporters. Physiology. 2007; 22:122–130.
Prassas, I, Diamandis, EP. Novel therapeutic applications of cardiac glycosides. Nat Rev Drug Discov. 2008; 7:926–935.
Seyberth, HW, Schlingmann, KP. Bartter- and Gitelman-like syndromes: salt-losing tubulopathies with loop or DCT defects. Pediatr Nephrol. 2011; 26:1789–1802.
Thorens, B, Mueckler, M. Glucose transporters in the 21st century. Am J Physiol Endocrinol Metab. 2010; 298:E141–E145.
Transport across cell membrane. http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/D/Diffusion
Membrane transpor. www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb1/part2/carriers.htm
Human ABC transporter. http://nutrigene.4t.com/humanabc.htm
P-ATPas. www.traplabs.dk/patbase