The Cervical Region
Characteristics of the Cervical Spine as a Whole
External Aspect of the Occipital Bone
Atypical and Unique Cervical Vertebrae
Atlas (First Cervical Vertebra)
Axis (Second Cervical Vertebra)
Carotid Tubercles and Articular Processes of the Sixth Cervical Vertebra
Articulations of the Upper Cervical Region
Clinical Applications Related to the Atlas and Axis
Ligaments of the Cervical Region
Ranges of Motion of the Cervical Spine
Nerves, Vessels, Anterior Neck Muscles, and Viscera of the Cervical Region
The cervical region is possibly the most distinct region of the spine. The fact that so many structures, spinal and otherwise, are packed into such a small cylinder, connecting the head to the thorax, makes the entire neck an outstanding feat of efficient design. The cervical spine is one of the most complicated articular systems in the body, comprising 76 separate joints (see Appendix 1). It allows more movement than any other spinal region and is surrounded by a myriad of nerves, vessels, and many other vital structures. However, such complexity can come at a high cost; more than 50% of individuals suffer from significant neck pain at some time in their lives (Jonsson, 2000, SBU Project Group). All clinicians who have spent significant time working with patients suffering from pain of cervical origin have been challenged and sometimes frustrated with this region of immense clinical importance. Understanding the detailed anatomy of this area helps clinicians make more accurate assessments of their patients, which in turn results in the establishment of more effective treatment protocols.
This chapter begins by covering the general characteristics of the cervical spine as a whole. This is followed by a discussion of the region’s typical and atypical vertebrae. The external aspect of the occiput is included because of its intimate relationship with the upper two cervical segments. Then the ligaments of the cervical region are covered, followed by a discussion of the cervical spine’s ranges of motion. The most important structures of the anterior neck and cervical viscera also are included.
Characteristics of the Cervical Spine as a Whole
The cervical curve is the least distinct of the spinal curves. It is convex anteriorly (lordosis) and is a secondary (compensatory) curvature (see Chapter 2). The cervical curve begins to develop before birth at as early as 9 weeks of prenatal life. Onset of fetal movements plays an important role in the early development of the cervical lordosis (Bagnall, Harris, & Jones, 1977; Standring et al., 2008). However, the curve becomes much more marked when the child begins to lift the head at approximately 3 to 4 months after birth, and the curve increases as the child begins to sit upright at approximately 9 months of age (Standring et al., 2008).
The amount of cervical lordosis can be measured reliably from standard x-rays if rigorous, standardized methods of evaluation are used (Côté et al., 1997). The cervical lordosis increases significantly with age in both sexes. In males, the change is rather gradual until the age of 70, when a dramatic increase in lordosis occurs. In females, in whom the cervical lordosis is greater throughout life, the curve increases gradually until a time of several years of marked increase begins near the age of 50; the rate of change then becomes more gradual but continues throughout the remainder of life (Doual, Ferri, & Laude, 1997).
Some authors state that the cervical curve actually is composed of two curves, upper and lower (Kapandji, 2008; Oliver & Middleditch, 1991). The upper cervical curve is described as a distinct primary curve that extends from the occiput to the axis and is concave anteriorly (kyphotic). The lower cervical curve is the classically described lordosis, but in this case begins at C2 rather than C1. This description helps to explain the dramatic differences seen between the upper and lower cervical vertebrae, such as the independent movements that can occur in the two regions (e.g., flexion of the lower cervicals and simultaneous extension of the occiput on the atlas and the atlas on the axis).
The lack of a normal cervical lordosis is a frequent finding in children and adolescents younger than age 17, but a lack of cervical lordosis in the adult may be a sign of ligamentous injury (Fesmire & Luten, 1989) or anterior cervical muscular hypertonicity. However, some controversy exists on this topic. Some authors (G. Schultz, personal communication) consider a hypolordotic or even kyphotic cervical curve to be clinically insignificant, especially if a tendency toward the establishment of a lordosis can be demonstrated during extension of the cervical region. Methods used to evaluate such a tendency include palpation at the spinous processes during extension, procurement of lateral projection x-ray films while the patient holds the neck in extension, and performance of cineradiography studies in which a series of x-ray films are taken while the patient moves the neck. Another cause of decreased cervical lordosis is the presence of hyperplastic articular pillars (see Articular Processes and Zygapophysial Joints). Hyperplastic articular pillars are considered to be a common normal variation of the typical cervical vertebrae (C3-6) and are associated with a reduction of the cervical lordosis (Peterson et al., 1999).
Total craniospinal height decreases significantly from early adulthood to senescence in both sexes, but the rate of change is different in males and females. In males, the change is more gradual, whereas in females, in whom the overall decrease in height is greater than that in males, there is a marked change near the age of 50, presumably associated with menopause. As described, the cervical lordosis also markedly increases during this time. The rate of decrease in height of the cervical region then slows again after the fifth decade (Doual, Ferri, & Laude, 1997).
Degenerative joint disease (DJD; bone spur or osteophyte formation, and intervertebral disc narrowing) occurs earlier in males (31 to 40 years of age) than females. However, the incidence of female DJD of the cervical region equalizes as a sharp rise in DJD occurs at 51 to 60 years of age. After 60 years of age the incidence between males and females is approximately equal (Wiegand et al., 2003).
Typical Cervical Vertebrae
The typical cervical vertebrae are C3 through C6. These are some of the smallest but most distinct vertebrae of any vertebral region. C1 and C2 are considered to be atypical vertebrae, and C7 is unique. These three vertebrae are discussed later in this chapter.
The individual components of the typical cervical vertebrae are discussed in the following section. Special emphasis is placed on those characteristics that distinguish typical cervical vertebrae from the other spinal vertebrae. Figure 5-1 illustrates typical cervical vertebrae and Figure 5-2 shows the entire cervical spine (including x-rays).
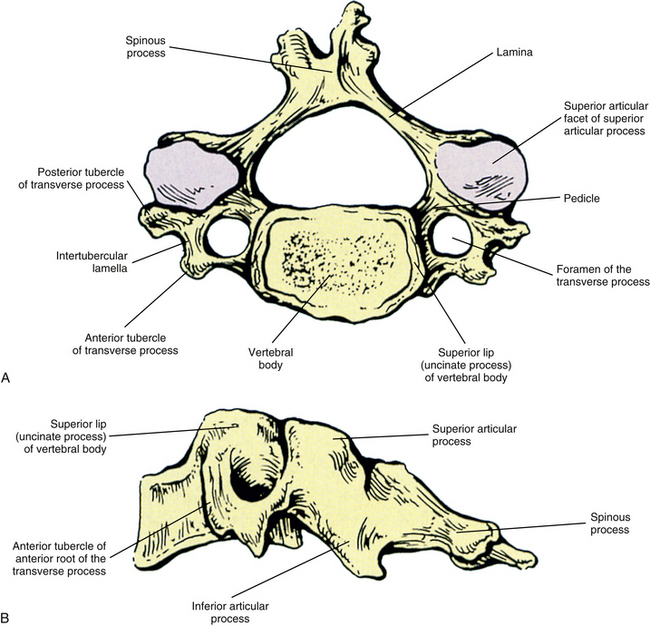
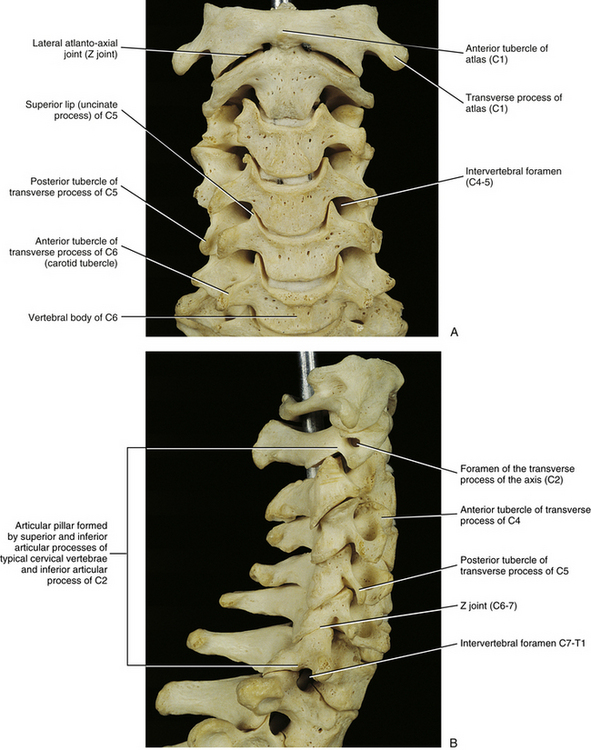
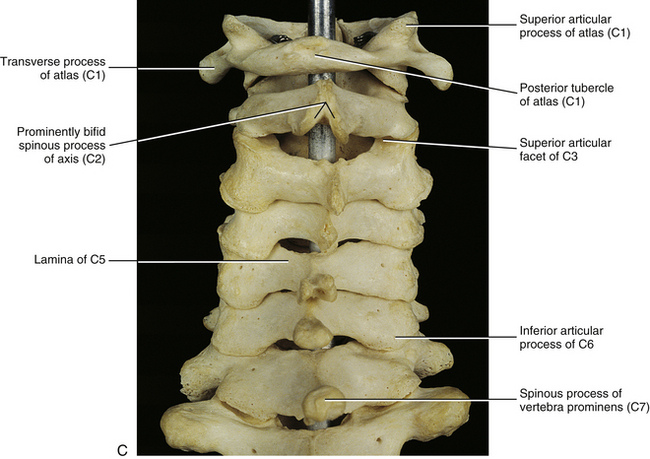
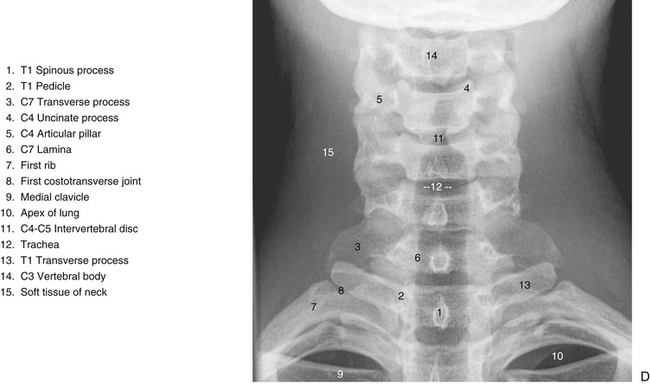
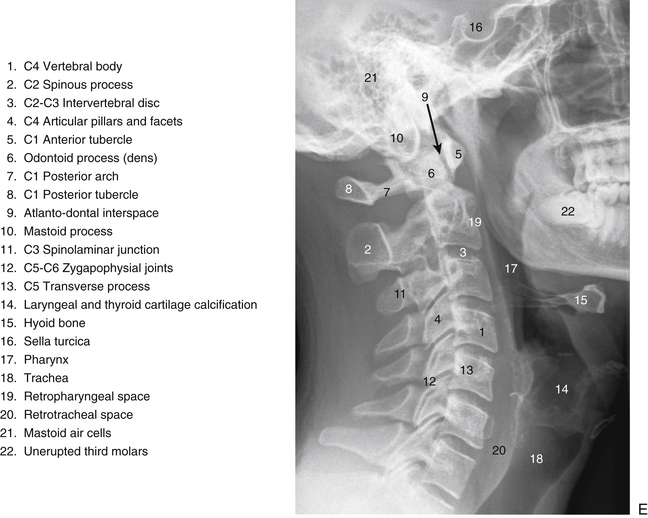
FIG. 5-2 A, Anterior, B, lateral, and, C, posterior views of the cervical region of the vertebral column. A, Superior lips (uncinate processes) of the C3 to C6 vertebral bodies are shown to advantage. B, Note the articular pillar, formed by the C3 through C6 superior and inferior articular processes. C, Posterior view of the cervical region of the vertebral column. D, Anterior-posterior lower cervical spine x-ray. E, Lateral cervical spine x-ray. (D and E, Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
Vertebral Bodies
Each cervical vertebra consists of a vertebral body and a posterior arch (Fig. 5-1). The vertebral bodies of the cervical spine are small and are more or less rectangular in shape when viewed from above, with the transverse (left-to-right) diameter greater than the anteroposterior diameter (transverse diameter 24.6 ± 2.4 and 23.0 ± 2.4 mm and anteroposterior diameter 17.1 ± 0.2 and 15.2 ± 0.3 mm in males and females, respectively). Because of the “inferior lip” of the anterior aspect of the inferior vertebral margin, the anteroposterior distance is greater at the inferior than the superior aspect of the vertebral bodies. The exception to this is C7, where the anteroposterior distance is approximately equal at the superior and inferior vertebral margins. The transverse and anterior-to-posterior diameters both increase from C2 to C7 with the transverse increasing more than the anteroposterior diameters (Kwon et al., 2004).
This allows the lower vertebrae to support the greater weights they are required to carry. In addition, the vertebral bodies of males have been found to be larger than those of females (Liguoro, Vandermeersch, & Guerin, 1994; Lu et al., 1999). The thickness of the outer cortex of the superior and inferior aspects of the vertebral bodies increases from C3 to C7. In addition, the superior aspect of each C3-7 vertebral body (superior bony end plate) is thickest posteriorly and thinnest anteriorly; the opposite is true for the inferior end plate (i.e., thickest anteriorly). The center of the inferior bony end plate also is thicker than the center of the superior bony end plate in the C3-7 vertebrae. Finally, the heights (superior to inferior dimensions) of both the anterior and posterior aspects of the cervical vertebral bodies generally increase from C3 to C7 (Panjabi et al., 2001a).
The anterior surface of the cervical vertebral body is convex from side to side. However, these same surfaces are concave from superior to inferior (Panjabi et al., 2001a) because of prominent ridges at the superior and inferior borders (discal margins) formed by the attachment sites of the anterior longitudinal ligaments. Indentations seen on the left and right of the anterior midline of the vertebral bodies are for attachment of the vertical fibers of the longus colli muscle.
The anterior aspects of the vertebral bodies can develop bony spurs (osteophytes). Asymptomatic osteophytes may occur in 20% to 30% of the population. Pressure on the more anteriorly located esophagus or trachea from osteophytes may lead to difficulty with swallowing (dysphagia) and speech (dysphonia), although such consequences are rare (Kissel & Youmans, 1992).
The posterior surface of a typical cervical vertebral body is slightly indented (Panjabi et al., 2001a) and possesses two or more foramina for exit of the basivertebral veins (Standring et al., 2008). The posterior longitudinal ligament attaches to the superior and inferior margins of the posterior aspect of the cervical vertebral bodies.
The superior and inferior surfaces of the vertebral bodies typically are described as being sellar or saddle shaped. More specifically, the superior surface is concave from left to right as a result of the raised lateral lips (uncinate processes). The superior surface is also convex from front to back because of the beveling of its anterior aspect. The inferior surface is convex from left to right and concave from anterior to posterior. Much of the concavity is created by the anterior lip of the inferior surface. This concavity is greatest at C3 and C4 and then becomes shallower as one moves inferiorly from C5 to C7 (Panjabi et al., 2001a). The inferior aspects of the vertebral bodies of C5 and C6 have the greatest anteroposterior dimension (Lu et al., 1999). Osteophyte (bone spur) formation is also most common at the C5-6 levels (Wiegand et al., 2003).
The anteroinferior aspect of each vertebral body usually protrudes inferiorly to overlap the anterior portion of the intervertebral disc (IVD) and occasionally the vertebra below. When the latter occurs, the anterosuperior aspect of the vertebra below is more beveled than would otherwise be the case. This increased beveling allows the vertebra to receive the projecting portion of the body above during flexion of the cervical spine.
Raised Lips at the Superior Aspect of the Vertebral Bodies and the Uncovertebral Joints
When viewed from the lateral or anterior aspect, several unique characteristics of the vertebral bodies become apparent (see Figs. 5-1, B, and 5-2, A). Lateral lips (uncinate processes) project from the superior surface of each typical cervical vertebra. These structures arise as elevations of the lateral and posterior rims on the top surface of the vertebral bodies (Dupuis et al., 1985). The posterior components of the uncinate processes tend to become more prominent in the lower cervical vertebrae. Normally the uncinate processes allow for flexion and extension of the cervical spine and help to limit lateral flexion. In addition, the uncinate processes serve as barriers to posterior and lateral IVD protrusion. The relationship of the left and right uncinate processes to the IVD has led at least one investigator to state that IVD herniation in the cervical region may occur less frequently than previously considered (Bland, 1989).
The uncinate processes of one vertebra may articulate with the small indentations found on the inferior surface of the vertebra above by means of small synovial joints. These joints are sometimes called the uncovertebral joints (of von Luschka). They consist of oblique clefts that develop at approximately 9 to 10 years of age. The clefts are limited medially by the IVD and laterally by capsular ligaments (Standring et al., 2008); the latter are derived from the anulus fibrosus of the IVD. Some investigators do not believe that the uncovertebral joints can be classified as synovial joints (Tondury, 1943; Orofino, Sherman, & Schecter, 1960; Bland, 1989), whereas others believe they do possess a synovial lining (Cave, Griffiths, & Whiteley, 1955). Regardless of their true classification, the distance between the apposing bony structures in the uncovertebral region is only approximately one third of that of the remainder of the interbody space (Hadley, 1957). In addition, the uncovertebral joints frequently undergo degeneration with resulting bony outgrowth (osteophyte formation) of the uncinate processes. These osteophytes may encroach on neighboring structures such as the vertebral artery and exiting cervical spinal nerves within the intervertebral foramen (Hadley, 1957; Bland, 1989). The uncinate processes are significantly higher at C4 to C6 (5.8 ± 1.1 to 6.1 ± 1.3 mm) than at C3 or C7 levels, and the length of the exiting dorsal and ventral roots and spinal nerve from the dura mater to the lateral border of the intervertebral foramina increases from C3 to C7. The higher uncinate processes at C4-6 and the longer nerves in this region may help to explain the higher incidence of neural compression syndromes associated with the C4-6 vertebral levels (Ebraheim et al., 1997c). Furthermore, dense fibrous tissue attaches to the uncinate process and this tissue then encases the spinal nerve and vertebral artery in the intertransverse space at the same level. The vertebral artery may become vulnerable to irritation, or in extreme cases perforation, from osteophytes projecting from the uncinate processes, because in essence the vertebral artery is held in a relatively close relationship to the uncinate process by this uncinate–spinal nerve–vertebral artery fibrous complex (Ebraheim et al., 1998; Yilmazlar et al., 2003a,b). Because the vertebral artery courses closer to the uncinate processes in the midcervical region than in the lower cervical region, the vertebral artery in the midcervical region is potentially more vulnerable than the vertebral artery in the lower cervical region to damage from osteophytes arising from the uncinate processes (Ebraheim et al., 1997c).
Injury to the vertebral bodies frequently, but not always, results in swelling of the prevertebral soft tissues (Miles & Finlay, 1988). Therefore swelling of the prevertebral tissues seen on standard x-ray films after trauma is an indication for further diagnostic studies such as computed tomography (CT).
Pedicles
The left and right pedicles of a typical cervical vertebra (see Fig. 5-1) are small and project posterolaterally from the vertebral bodies at approximately a 60-degree angle to the frontal (coronal) plane. They form the medial boundary of the left and right foramina of the transverse processes, respectively. The width (4.4 to 4.9 mm), height (6.7 to 7.2 mm), and length (5.3 to 6.2 mm) of the pedicles are relatively similar in the C3 to C7 vertebrae (Bozbuga et al., 2004), but the pedicles of C2 are shorter than those of the rest of the cervical region (Karaikovic et al., 1997). The pedicles of typical cervical vertebrae are placed about midway between the superior and inferior margins of the vertebral body. Therefore the superior and inferior vertebral notches are of approximately equal size (Standring et al., 2008). The spinal nerve, surrounded by a sleeve of dura, courses just above (superior to) the pedicle of each typical cervical vertebra.
The cervical pedicles have a relatively thick layer of cortical bone (also known as compact bone) that surrounds a central core of cancellous bone (also known as trabecular, or spongy, bone). Generally, the cortex of the lateral aspect of the pedicle (the region closest to the vertebral artery) is thinner than that of the medial side. However, approximately 4% of pedicles are composed entirely of cortical bone with no medullary cavity. In the remainder of pedicles, the central trabecular bone is continuous with that of the more posteriorly located articular processes. This allows transfer of loads from the vertebral body to the articular pillar (discussed later in this chapter) during flexion and from the articular pillar to the vertebral body during extension (Pal et al., 1988).
Transverse Processes
The left and right transverse processes (TPs) of a typical cervical vertebra are each composed of two roots, or bars, one anterior and one posterior (see Fig. 5-1). The two roots end laterally as tubercles (anterior and posterior). Occasionally an anterior tubercle of one vertebra is significantly longer than those of its neighbors. This usually occurs at C5 or C6 and has been associated with a lack of segmentation (congenital fusion) of the vertebral body of the segment with the long anterior tubercle to the vertebral body directly above or below (Ehara, 1996). The two tubercles are joined to one another by an intertubercular lamella, which is less correctly known as a costotransverse lamella (bar) (Standring et al., 2008). The distance between the lateral tips of the left and right TPs is greatest at C1. This same distance, although smaller, remains relatively constant from C2 through C6 and then increases greatly at C7.
In the typical cervical vertebrae a gutter, or groove, for the spinal nerve is formed between the anterior and posterior roots of each TP (Fig. 5-3). This groove serves as a passage for exit of the spinal nerve and its largest branch, the anterior primary division (ventral ramus) (Fig. 5-3, B). The neural grooves form approximately a 50-degree anterior angle with the midsagittal plane, except for C7 where the angle is larger, usually 56 to 57 degrees. Generally, there is no significant difference between the length of the neural grooves in males and females. The depth of the neural grooves gradually increases from C3 (≈3 ± 1.0 mm deep) to C7 (≈5 ± 0.7 mm deep) (Ebraheim, Biyani, & Salpietro, 1996). The intertubercular lamellae of C3 and C4 have an oblique course, descending from the anterior root and passing laterally as they reach the posterior root of the TP. Therefore the anterior tubercles (roots) of these vertebrae are shorter than the posterior ones, and the grooves for the spinal nerves are deeper posteriorly than anteriorly. The left and right intertubercular lamellae of C6 are wide (from left to right) and shallow (from superior to inferior) (Standring et al., 2008).
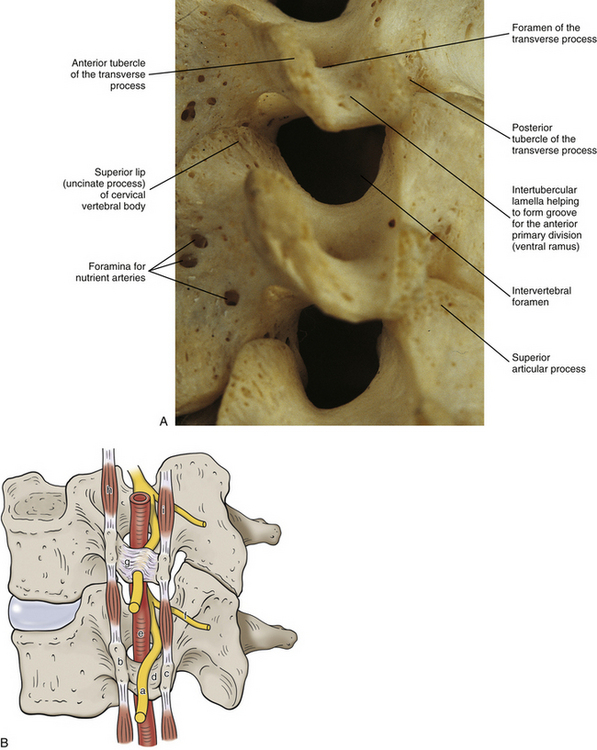
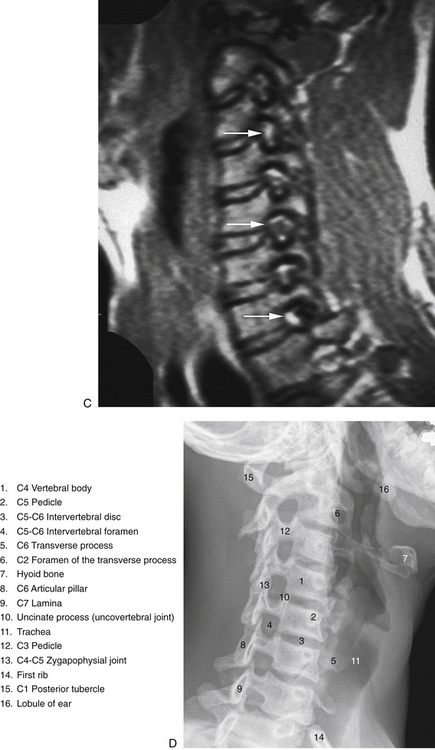
FIG. 5-3 Obliquely oriented cervical intervertebral foramina (IVFs). A, Close-up of several cervical IVFs. Notice that the superior lip (uncinate process) of a typical cervical vertebral body helps to form the anterior border of the IVF. B, Groove, or “gutter,” for the spinal nerve (GSN) formed by the anterior and posterior tubercles and intertubercular lamella of each transverse process of typical cervical vertebrae. Notice that the groove for the spinal nerve is actually the groove for the anterior primary division (APD). Also, notice the normal connective tissue holding the APD in place has been illustrated in the GSN of the upper transverse process. This connective tissue frequently extends proximally to the spinal nerve. The connective tissue protects the vulnerable cervical dorsal and ventral roots from traction injuries that could be caused by extreme movements of the neck or upper extremities. The connective tissue is removed in the lower transverse process to show the course of the APD along the intertubercular lamella. The following structures are identified: a, anterior primary division (ventral ramus); b, anterior tubercle (of transverse process); c, posterior tubercle; d, intertubercular lamella; e, vertebral artery; f, posterior primary division (dorsal ramus); g, connective tissue surrounding the APD and holding the APD to the groove for the spinal nerve; h, anterior intertransversarius m.; i, posterior intertransversarius m. C, Because the cervical IVFs face anteriorly as well as laterally, MRI scans taken at a 40- to 45-degree angle to a sagittal plane show the cervical IVFs to best advantage. Notice that in this direct oblique sagittal MRI the C2-3 through T1-2 IVFs all can be seen (arrows are pointing to the C3-4, C5-6, and C7-T1 IVFs). D, Oblique x-ray of the cervical region. (C, Courtesy Dr. Dennis Skogsbergh. D, From Cramer G et al. [2002]. Oblique MRI of the cervical intervertebral foramina: a comparison of three techniques. J Neuromusculoskel Syst, 10, 41-51. E, Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
A dural root sleeve, which surrounds each spinal nerve, and also its continuation as the epineurium of the ventral ramus (anterior primary division) are held to the gutter of the TP by fibrous tissue. This strong attachment to the TP is unique to the cervical region. More specifically, the C4, C5, and C6 spinal nerves and anterior primary divisions (ventral rami) are held in place to the groove for the spinal nerve by means of tough connective tissue attachments (Fig. 5-3, B). The nerves at the other levels are not as tightly bound to the bone of the groove for the spinal nerve. Lateral traction of the dura (more specifically the “dural funnel”) pulls it laterally into an intervertebral foramen (IVF) until it plugs the foramen. This prevents further traction on the nerve roots and rootlets and their attachment to the spinal cord. In addition, traction of nerves also tractions the denticulate ligament, which in turn causes the spinal cord to move laterally toward the traction force, thus removing tension on the spinal cord from the nerve roots undergoing traction (Sunderland, 1974).
The dural root sleeves in the cervical region receive more sensory innervation than any other region of cervical dura, and the fibers innervating the dural root sleeves originate directly from the dorsal root ganglia, not the recurrent meningeal nerves (Yamada et al., 1998). This means significant traction or compression of the cervical dural root sleeves could result in the production of pain.
A dorsal ramus leaves each spinal nerve shortly after the spinal nerve is formed by the union of the dorsal and ventral roots. The dorsal ramus (posterior primary division) courses posteriorly and laterally along the zygapophysial joint (see next section), supplying the joint with sensory innervation. The ramus then passes posteriorly to supply the cervical parts of the deep back muscles with motor, nociceptive, and proprioceptive innervation; and then continues posteriorly to reach the dermal and epidermal layers of the back to supply them with sensory innervation. The nerves of the cervical region are discussed in more detail later in this chapter.
The anterior aspects of the TPs of C4 to C6 terminate in roughened tubercles that serve as attachments for the tendons of the scalenus anterior, longus colli (superior and inferior oblique fibers), and longus capitis muscles. The posterior tubercles extend further laterally and slightly more inferiorly than their anterior counterparts (except for C6, where they are level). The splenius cervicis, longissimus cervicis, iliocostalis cervicis, levator scapulae, and scalenus medius and posterior muscles attach to the posterior tubercles.
As the name implies, the foramen of the TP is an opening within the TP. This foramen is present in the left and right TPs of all cervical vertebrae. It was previously called the foramen transversarium, but the currently preferred term is simply foramen of the transverse process. The boundaries of this foramen are formed by four structures: the pedicle, anterior root of the TP, posterior root of the TP, and intertubercular lamella. The sizes of left and right foramina of the TPs of a single vertebra frequently are asymmetric, and occasionally the foramen of a single TP is double (Taitz, Nathan, & Arensburg, 1978). The vertebral artery normally enters the foramen of the TP of C6 and continues superiorly through the corresponding foramina of C5 through C1. The vertebral artery of each side loops posteriorly and then medially around the superior articular process of the atlas on the corresponding side. The artery then continues superiorly to pass through the foramen magnum. The ventral rami of the C3 to C6 spinal nerves pass directly posterior to the vertebral artery. In fact, each of these rami usually rests against the artery as the ventral ramus exits the gutter (groove) for the spinal nerve of the TP (Paolini & Lanzino, 2006) (see Fig. 5-3).
Several vertebral veins on each side also pass through the foramina of the TPs. These veins begin in the atlanto-occipital region and continue inferiorly through the foramina of the TPs of C1 through C7 and then enter the subclavian vein. The vertebral veins receive branches from both the epidural venous plexus and the external vertebral venous plexus. In addition to the veins, a plexus of sympathetic nerves also accompanies the vertebral artery as it passes through the foramina of the TPs of C1 through C6 (see Fig. 5-32). The vertebral artery and sympathetic plexus associated with it are discussed in more detail later in this chapter.
Cervical ribs: As mentioned in Chapter 2, the vertebrae of each region of the spine possess specific sites that are capable of developing ribs. Such regions are known as costal elements, costal processes, or pleurapophyses. The cervical region is no exception. The costal process of a typical cervical vertebra constitutes the majority of its TP. In fact, all but the most medial aspect of the posterior root of the TP participates in the formation of the costal process. The costal processes may develop into cervical ribs in some individuals (Standring et al., 2008). This occurs most frequently at the level of C7. A cervical rib at C7 may compress underlying portions of the brachial plexus and the subclavian artery. The symptom complex that results from compression of these structures is commonly known as thoracic outlet syndrome, and a cervical rib is one cause of this syndrome (Bland, 1987). (Thoracic outlet syndrome is discussed in detail in Chapter 6.) A cervical rib may develop as a small projection of the TP or may be a complete rib that attaches to the manubrium of the sternum or the first thoracic rib. However, the cervical rib usually is incomplete, and a bridge of fibrous tissue usually connects the tip of the cervical rib to either the manubrium or the first thoracic rib. The osseous extension of the cervical TP frequently can be detected on standard x-ray films, but the fibrous band is much more difficult to evaluate radiographically.
Articular Processes and Zygapophysial Joints
The general characteristics of the articular processes and the zygapophysial joints (Z joints) are discussed in Chapter 2. The unique characteristics of the cervical Z joints are discussed here. The superior articular processes and their hyaline cartilage–lined facets face posteriorly, superiorly, and slightly medially (see Fig. 5-1), and the cervical Z joints lie approximately 45 degrees to the horizontal plane (White & Panjabi, 1990; Panjabi et al., 1991) (Figs. 5-1, 5-2, B, and 5-4). More specifically, the facet joints of the upper cervical spine lie at approximately a 35-degree angle to the horizontal plane, and the lower cervical Z joints form a 65-degree angle to the horizontal plane (Oliver & Middleditch, 1991). The articular processes and their hyaline cartilage–lined facets of the typical cervical vertebrae are more horizontally oriented at birth (averaging approximately 34 degrees to the horizontal plane) than in the adult. The articular processes then gradually become more vertically oriented until approximately 10 years of age, reaching an angle averaging approximately 45 to 53 degrees. They maintain this angle throughout the remainder of life. The seventh cervical vertebra follows the same pattern, but throughout life is more vertically oriented than the typical vertebrae. The more horizontally oriented articular processes of children (when compared with adults) lead to an increased anterior-to-posterior translation among the typical cervical vertebrae (Kasai et al., 1996).
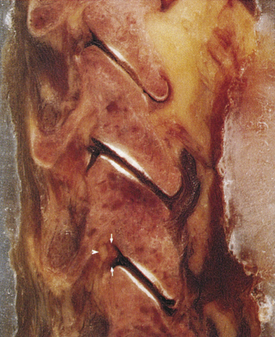
FIG. 5-4 Parasagittal section through the cervical Z joints. Anterior is left and posterior is right. Notice the white articular cartilage of the superior and inferior articular facets. The arrows indicate a small Z joint synovial fold. (From Yu S et al. [1987]. Facet joint menisci of the cervical spine: correlative MR imaging and cryomicrotomy study. Radiology, 164, 79-82.)
Right-to-left asymmetry of the plane of articulation of the cervical articular processes is common. This is particularly true at the junction of the cervical and thoracic regions. An asymmetry of greater than 10 degrees in the coronal (frontal) plane occurs 24% of the time at C6, 10% of the time at C7, and 16% of the time at T1. There is no significant difference in asymmetry of the spatial orientation of the articular processes between men and women (Boyle, Singer, & Milne, 1996).
The heights (anterior-to-posterior dimension) of the superior articular facets of C3-7 range from 10.0 to 8.7 mm, decreasing in height from C3 to C7. The widths are approximately 11.0 mm from C3 to C6 and increase to 12.3 mm at C7. The height and width of the superior articular processes of males are again greater than those of females (Ebraheim et al., 1997b).
The heights (anterior-to-posterior dimension) of the inferior articular facets range from 9.0 to 9.6 mm from C3 to C7; C3 has the largest height and C5 the smallest. There is a steady decrease in height from C3 to C5; then a slight increase at C6 and again at C7. Therefore unlike the superior articular processes, there is no steady trend in height of the inferior articular processes. The width of the inferior articular processes ranges from 11.2 to 12.1 mm, and there is a small, steady increase from C3 to C7. The height and width of the inferior articular processes of males are greater than those of females (Ebraheim et al., 1997b).
The appearance of the cervical Z joints changes significantly with age. Before age 20 the articular cartilage is smooth and approximately 1.0 to 1.3 mm thick, and the subarticular bone is regular in thickness. The articular cartilage thins with age, and most adult cervical articular facets possess an extremely thin layer of cartilage supported by irregularly thickened subarticular cortical bone. Articular cartilage degenerative changes include “splits” of the cartilage parallel to the joint surface; “fissures,” which are cartilage cracks perpendicular to the joint surface; and “flaking” of the cartilage adjacent to the joint space. The cartilage changes can begin at a rather young age and males show more cartilage changes than females. The subcortical bone thickens with age, approximately equally in both sexes (Uhrenholt et al., 2008). Such changes of articular cartilage and the subchondral bone usually are undetected on CT and magnetic resonance imaging (MRI) scans. Osteophytes (bony spurs) projecting from the articular processes and sclerosis (thickening) of the bone within the articular processes occur often in adult cervical Z joints (Fletcher et al., 1990; Kettler, Werner, & Wilke, 2007; Uhrenholt et al., 2008). Osteophytes of the superior articular processes most frequently extend from the anterior surface (17% of spines) into the region of the intervertebral foramen (Kettler, Werner, & Wilke, 2007) and may compress the neural elements coursing through this region (Hackney, 1992).
The Z joint capsules probably do little to limit normal motion in the cervical region (Onan, Heggeness, & Hipp, 1998); however, the anterior aspects of the capsules do restrain hyperextension (Samartzis et al., 2010). They are thicker along the lateral aspect of the joint compared with the posterior aspect, and become even thicker from posterior to anterior along the lateral aspect of the capsule, making the anterolateral aspect the thickest region of the Z joint capsule (Tonetti et al., 1999). However, overall the Z joint articular capsules of the cervical region are thin (Panjabi et al., 1991) and are longer and looser than those of the thoracic and lumbar regions. The collagen fibers that comprise the capsules course from the region immediately surrounding the articular facet of the inferior articular process of the vertebra above to the corresponding region of the superior articular process of the vertebra below (Fig. 5-2, B; see also Figs. 5-17 and 5-24). The bands of collagen fibers are approximately 9 mm long and course perpendicular to the plane created by the Z joint (Panjabi et al., 1991).
Two (one arising from the anterior pole and the other from the posterior pole) Z joint synovial folds (menisci) of approximately equal size project 1 to 5 mm into the Z joints at all levels of the cervical spine, and together the anterior and posterior synovial folds cover approximately one third of the articular surface (Mercer & Bogduk, 1993; Uhrenholt et al., 2008). The folds are present in both children and adults, although the synovial folds of children may be relatively larger and are less fibrous than those of adults (Tang et al., 2007). Smaller and more evenly distributed connective tissue (capsular) rims are sometimes found along the peripheral aspects of Z joints, but do not normally reach the articular cartilage; they have been found at all cervical levels (Mercer & Bogduk, 1993).
The synovial folds at the C1-C2 Z joint are larger than those of the other cervical Z joints, covering a higher percentage of the articular cartilage (Yu, Sether, & Haughton, 1987). The left and right anterior (anterolateral) C1-C2 folds are generally larger than the posterior folds (Tang et al., 2007; Webb et al., 2009, 2011). The C1-C2 synovial folds have been implicated as a source of neck pain and headache following the flexion-extension injuries (i.e., whiplash-associated disorders) of automobile accidents, because postmortem pathologic changes have been found in these folds in victims of automobile accidents (Webb et al., 2009). The C1-C2 synovial folds have been imaged with clinically applicable MRI protocols (Webb et al., 2009), whereas the synovial folds of the lower cervical Z joints have been visualized on MRI scans under experimental conditions (Yu, Sether, & Haughton, 1987; Friedrich et al., 2007) and with 3 Tesla (high-field strength) MRI units (Friedrich et al., 2008). Figure 5-3 shows a small Z joint synovial fold.
The proposed normal functions of the Z joint synovial folds include lubricating the Z joints, acting as passive space fillers in the Z joints, and possibly serving joint-stabilizing and force-dissipating roles (Mercer & Bogduk, 1993; Webb et al., 2011). As mentioned previously, damage has been identified in these folds following whiplash-type injuries. Because the synovial folds have a sensory innervation such damage could be a source of neck pain and headache in whiplash patients. In addition, other hypotheses have been forwarded implicating the synovial folds in back (see Chapters 6 and 7) and neck pain. One such theory is that the synovial folds become trapped outside of the joint space, between an articular process and the Z joint capsule. This has been called the extrapment theory; however, extrapment of Z joint synovial folds is currently thought to occur more often in the thoracic region than in the cervical region (Schulte et al., 2010; Webb et al., 2011). Another hypothesis that appears to have the most biologic plausibility in the cervical region is known as the entrapment theory. Entrapment refers to the Z joint synovial folds becoming “pinched,” or trapped, within Z joints when the folds fail to move peripherally, as usually occurs when compressed during joint motion (Kos, Hert, & Sevcik, 2002; Webb et al., 2011). Entrapped synovial folds have been implicated as a possible cause of torticollis in some patients (Mercer & Bogduk, 1993). Entrapped synovial folds have been visualized in the posterior aspect of the atlanto-occipital joint in a very low percentage (0.36%) of healthy individuals (Friedrich et al., 2008); however, further advances in imaging are needed to routinely assess Z joint synovial folds. The folds may also form a nidus for the development of intraarticular Z joint adhesions (Mercer & Bogduk, 1993). Chapter 7 discusses the synovial folds in the lumbar region and Chapter 14 (Figs. 14-1 and 14-7) discusses the histologic composition of the folds.
When the individual vertebrae are united, the articular processes of each side of the cervical spine form an articular pillar that bulges laterally at the pediculolaminar junction of each vertebra (Standring et al., 2008). This pillar is conspicuous on lateral x-ray films. The cervical articular pillars (left and right) help to support the weight of the head and neck (Pal et al., 1988). Therefore weight bearing in the cervical region is carried out by a series of three longitudinal columns: one anterior column, which runs through the vertebral bodies; and two posterior columns, which run through the right and left articular pillars (Louis, 1985; Pal et al., 1988). Consequently, loss of intervertebral disc height (e.g., disc height loss as a result of intervertebral disc degeneration) usually results in increased loads being placed on the cervical articular facets (Hussain et al., 2010a,b).
Articular pillar fracture is fairly common in the cervical spine and frequently is undetected (Renaudin & Snyder, 1978). This type of fracture is usually a chip fracture of a superior articular facet and process. The patient often experiences transient radicular pain (see Chapter 11), which usually is followed by mild to intense neck pain. Persistent radiculopathy in such patients indicates displacement of the fractured facet and articular process onto the dorsal root within the intervertebral foramen (Czervionke et al., 1988) (see Fig. 5-5).
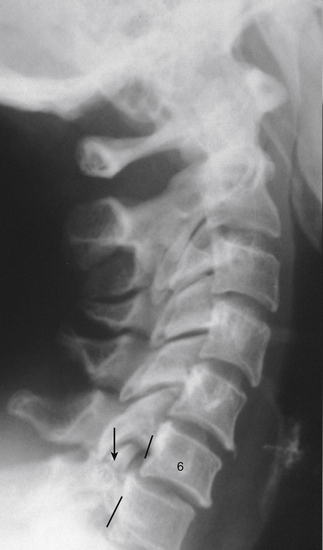
FIG. 5-5 Lateral cervical spine x-ray showing a traumatic anterolisthesis of C6 (6) secondary to fracture of the superior articular process of C7 (arrow). (Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
Spondylolysis of a cervical vertebra is defined as a complete cleft between the superior and inferior articular processes—the region equivalent to the pars interarticularis of a lumbar vertebra. Spondylolysis can be unilateral or bilateral. Unlike the lumbar region where the condition is common and generally thought to be a type of fracture, spondylolysis in the cervical region is very rare (the cervical articular pillars are quite substantial) and there is no consensus as to whether the offending cleft in the cervical vertebra is congenital in origin or is the result of repeated “microtrauma.” Spondylolysis has been identified in all cervical vertebrae with the exception of the atlas (C1). The vertebra most commonly affected is C6, with C4 and C5 being next in incidence (Paik, 2010). Spondylolysis is most common at the L5 vertebral level; consequently, the subsection entitled Spondylolysis and Spondylolisthesis of the section Fifth Lumbar Vertebrae in Chapter 7 describes this condition in detail.
The medial branch of the posterior primary division (dorsal ramus) of the spinal nerve provides sensory innervation by means of both mechanoreceptors and free nerve endings to the Z joint capsule. The cervical Z joints receive innervation from medial branches of two adjacent spinal segments, with the exception of the C2-C3 Z joint, which receives innervation from only the third occipital nerve (through a communication to the C2 posterior primary division) (Bogduk, 2005). This indicates that the Z joint capsules are important for providing information on both joint position and tissue damage to the central nervous system (McLain, 1994). The Z joints are a source of pain in a substantial number of both acute and chronic neck pain patients (Aprill & Bogduk, 1992), and are estimated to be a source of pain in approximately 50% of those with chronic neck pain (Bogduk, 2005). Pain arising from pathologic conditions or dysfunction of the cervical Z joints can refer to regions distant from the affected joint (Bogduk, 1989b; Aprill, Dwyer, & Bogduk, 1990; Dwyer, Aprill, & Bogduk, 1990). The two most common types of pain referral are neck pain and head pain (headache) arising from the C2-3 Z joints, and neck pain and shoulder pain arising from the C5-6 Z joints (Bogduk & Marsland, 1988). However, other Z joints (C1-C2 and C3-C4) have been implicated in some headaches (Webb et al., 2009; Lang & Buchfelder, 2010).
Laminae
The laminae of the cervical region are fairly narrow from superior to inferior. Therefore a gap can be seen between the laminae of adjacent vertebrae in a dried specimen (see Fig. 5-2, C). However, this gap is filled by the ligamentum flavum in live persons (see Fig. 7-20). The upper border of each cervical lamina is thin, and the anterior surface of the inferior border is roughened by the attachment of the ligamentum flavum. The ligamentum flavum is discussed in detail later in this chapter.
Vertebral Canal
A vertebral foramen of a typical cervical vertebra is triangular (trefoil) in shape (see Figs. 2-16, 5-1, and 5-6). It is also large, allowing it to accommodate the cervical enlargement of the spinal cord.
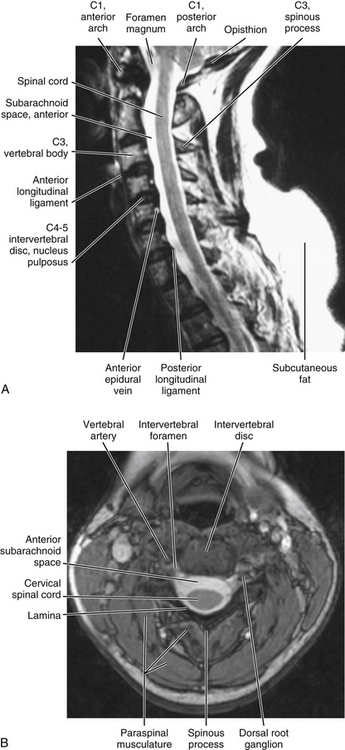
FIG. 5-6 Midsagittal (A, T2-weighted) and horizontal (axial) (B, gradient echo) MRIs showing the cervical vertebral canal and surrounding structures. Cerebrospinal fluid in the cervical subarachnoid space appears as high signal (white) on these images. (A and B, Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
Recall that the collection of all the vertebral foramina is known as the vertebral (spinal) canal. Therefore the IVDs and ligamenta flava also participate in the formation of the vertebral canal. The internal vertebral venous plexus lies within the epidural adipose tissue of the vertebral canal. The left and right longitudinal channels of the cervical anterior internal vertebral venous plexus are kept in the most anterior and lateral regions of the epidural space by a thin fibrous membrane on each side of the vertebral canal. These venous channels anastomose with one another behind each vertebral body via retrocorporeal veins that course between the posterior longitudinal ligament and vertebral bodies (Chaynes et al., 1998). The anterior internal vertebral veins of the upper cervical region (periodontal veins) have communications with the venous drainage of retropharyngeal structures via pharyngovertebral veins. These anastomoses have been implicated as a route for the spread of infection from anterior paravertebral structures to the occipital condyles and the atlas, in some cases leading to osteomyelitis affecting the upper cervical joints (Menezes & Traynelis, 2008).
The vertebral canal is fairly large in the upper cervical region but narrows from C3 to C6. More specifically, the transverse (left-to-right) dimension remains relatively constant at approximately 24.6 mm. However, the sagittal dimension becomes smaller; therefore the shape of the trefoil becomes relatively wider as one descends the vertebral canal in the cervical region (Hackney, 1992). In fact, the spinal cord occupies 75% of the space available within the vertebral canal at the C6 level. The volume of cerebrospinal fluid in the subarachnoid space in the cervical vertebral canal is least in extension and greatest in flexion, and the total change in volume of the cervical subarachnoid space (C2 to C7 levels) during full extension to full flexion is approximately 1.9 ml (Holmes et al., 1996). Table 5-1 summarizes the general characteristics of the cervical vertebral canal.
Table 5-1
General Characteristics of the Cervical Vertebral Canal
| Region | Dimensions |
| Upper cervical vertebral canal | Upper canal is infundibular in shape, wider superiorly than inferiorly; less than half the available space is occupied by the spinal cord at C1 |
| C4 | Narrowing of vertebral canal begins |
| C6 | Cord occupies 75% of vertebral canal |
| C1-7 | Critical anteroposterior dimension is 12-13 mm |
Connective tissue attachments to the posterior spinal dura mater: Connective tissue attachments to the posterior aspect of the spinal dura arising from the foramen magnum, posterior arch of C1, spinous process of C2 (Von Lanz, 1929), rectus capitis posterior minor muscle (Hack et al., 1995a,b; Zumpano, Jagos, & Hartwell-Ford, 2002), ligamentum nuchae (Mitchell, Humphreys, & O’Sullivan, 1998; Humphreys et al., 2003), and ligamenta flava between C1-2 and C6-7 (Shinomiya et al., 1995, 1996) have been described. These attachments may hold the dura mater posteriorly during cervical extension (to prevent buckling of the dura mater into the spinal cord) and flexion (to prevent the dura from moving forward and compressing the cord). Some authors have speculated that increased tension of the cervical paraspinal muscles may traction the connection between the rectus capitis posterior minor muscle and the dura, leading to headaches secondary to dural tension (Alix & Bates, 1999). Others have proposed that tearing of these connective tissue attachments during the flexion component of flexion-extension (whiplash) type of injuries or other trauma to the cervical region could lead to buckling of the dura mater into the cervical segments of the spinal cord. Such dural buckling could potentially result in chronic neck pain, headaches, disorders of balance, and signs and symptoms of cervical myelopathy experienced by some patients who have experienced trauma to the cervical region (Mitchell, Humphreys, & O’Sullivan, 1998). However, these theories remain conjecture until further basic science and clinical studies are performed to investigate the relationships between connective tissue attachments of the spinal dura mater and clinical conditions. In this regard, Shinomiya and colleagues (1995, 1996) have identified connective tissue attachments between the ligamenta flava at C1-2 and C6-7, which they call epidural ligaments. Using MRI scans of humans and spinal cord blood flow measurements and electrophysiologic recordings of the nerves innervating upper extremity muscles and from the spinal cord in an animal model (cat), they have identified a relationship between tearing or absence of the epidural ligaments at C6-7 and Hirayama-type amyotrophy. This condition is characterized by isolated muscle paralysis and atrophy of hand muscles resulting from segmental degenerative changes of the spinal cord gray matter. The authors found that absence or tearing of the epidural ligaments caused the spinal dura to press against the posterior aspect of the spinal cord during flexion of the spine. This compressed the spinal cord and its arterial supply, and led to a reduced blood flow to the anterior aspect of the spinal cord. The decreased blood flow to the spinal cord subsequently decreased motor activity of peripheral nerves to the upper extremity, and also decreased neuronal activity in the spinal cord at the C7 and C8 spinal cord segments. Their work provides strong evidence in support of an important physiologic role of connective tissue attachments to the posterior aspect of the spinal dura mater.
Cervical myelopathy: Pathology of the cervical spinal cord is termed cervical myelopathy. The development of bone spurs (osteophytes) both from the posterior aspect of the vertebral bodies and at articular processes close to the Z joints and uncinate processes has long been accepted as a potential cause of compression of the spinal cord and spinal nerve roots, respectively (Bailey & Casamajor, 1911; Boulos & Lovely, 1996). Other potential causes of spinal cord and nerve root compression include IVD protrusion, spinal cord tumor, Z joint hypertrophy, ossification of the posterior longitudinal ligament, buckling of the ligamentum flavum in a congenitally narrow vertebral canal, and a displaced fracture of a lamina, pedicle, or vertebral body. The critical anteroposterior dimension of the cervical vertebral canal is approximately 12 to 13 mm before manifestation of symptoms. A vertebral canal this narrow usually is the result of one of the aforementioned pathologic conditions combined with a congenitally narrow canal. However, the minimum dimension before symptoms occur may be slightly lower in blacks than in whites. The transverse and midsagittal diameters of the cervical vertebral canal are significantly less in black than white South Africans, although determining if the difference in dimensions results from heredity, environment, or prenatal and childhood nutrition is difficult. However, white South Africans have more bone spurs (spondylosis or osteophyte formation) affecting the size of the vertebral canal than black South Africans (Taitz, 1996).
When increased bone formation (spondylosis) of the vertebral bodies, uncinate processes, or the articular processes contributes to cervical myelopathy, the term cervical spondylotic myelopathy (CSM) is appropriate (Fig. 5-7). CSM develops slowly, is progressive, and also may be related to a congenitally narrow vertebral canal (Crawford, Cassidy, & Burns, 1995; Hukuda et al., 1996; McCormack & Weinstein, 1996). Less common causes of the condition are ossification of the posterior longitudinal ligament, ossification of the ligamentum flavum, and development of calcium pyrophosphate dehydrate deposition disease (Omura et al., 1996). In addition, Hukuda and colleagues (1996) found that large vertebral bodies (an increase of both the transverse and the sagittal diameters) were significantly related to CSM. They assumed this was because the large vertebral bodies resulted in larger osteophytes and larger disc bulges, therefore increasing the chances of clinically significant narrowing of the vertebral canal that would result in compression of the spinal cord. Cervical myelopathy in elderly persons frequently results from anterior or posterior translation of adjacent vertebrae (anterolisthesis or retrolisthesis), secondary to narrowing of the IVDs or injury to the cervical region (Kawaguchi et al., 2003). The chronic narrowing of CSM may reduce blood flow to the compressed spinal cord segments (Kurokawa et al., 2011). In addition, inflammation, initiated by arachnoid cells, may occur in the arachnoid mater in certain cases of CSM (Frank, 1995).
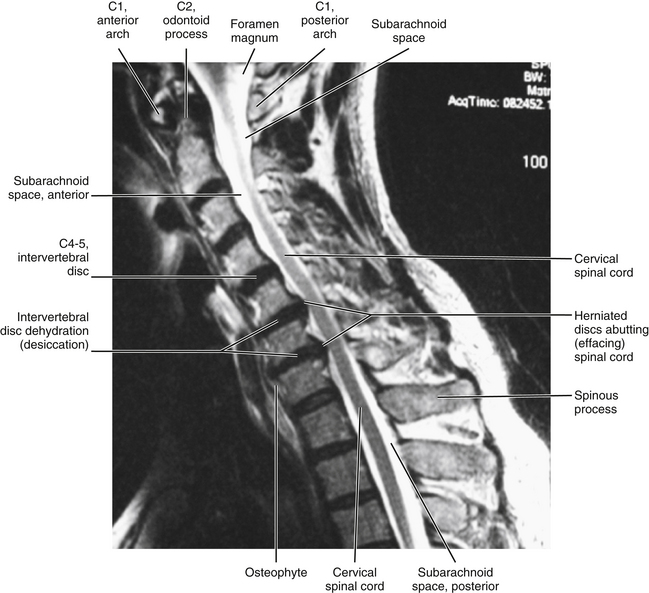
FIG. 5-7 Midsagittal MRI showing a case of cervical spondylotic myelopathy (CSM). Spondylosis (osteophytes) at C5-C6 and C6-C7 with posterior disc herniation extending to (“effacing”) the adjacent spinal cord. There is no evidence of spinal cord edema or spinal cord damage (e.g., myelomalacia, which is spinal cord softening that can occur with acute injury or severe intervertebral disc protrusion). Cerebrospinal fluid in the cervical subarachnoid space appears as high signal (white) on this T2-weighted image. (Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
The progression of spinal cord changes in CSM begins with neuronal loss in the anterior gray horn. This progresses to the intermediate gray matter. Degeneration then occurs in the lateral and posterior funiculi of white matter. The changes in the white matter are consistent with demyelination and remyelination similar to that which occurs in compression neuropathies of peripheral nerves, such as carpal tunnel syndrome. Finally, atrophy is seen throughout the entire gray matter and more severe degeneration is found in the lateral funiculus (Ito et al., 1996). These changes lead to decreased processing of afferent (sensory) input and efferent (motor) output of the affected spinal cord levels (Chistyakov et al., 1995).
CSM usually is associated with diffuse neck pain accompanied by varying degrees of neurologic deficit and can mimic multiple sclerosis, amyotrophic lateral sclerosis, intervertebral disc protrusion, and less commonly spinal cord tumor and subacute combined degeneration of the spinal cord (Cusick, 1988; Crawford, Cassidy, & Burns, 1995).
Vertebral canal measurements taken from the level of the superior border of the pedicle on standard lateral x-rays can be done reliably, and can be used before more advanced imaging procedures are employed to evaluate the size of the vertebral canal (Senol et al., 2001). Metrizamide myelography (injection of radiopaque dye followed by x-ray examination) and computer-assisted myelography (injection of dye followed by CT scanning) have been shown to be useful in the evaluation of cervical spondylotic myelopathy (Yu, Sether, & Haughton, 1987). Measurement (morphometry) of the cervical cord by means of MRI has been shown to correlate well with the severity of cord compression (Fujiwara et al., 1988).
Neurapraxia of cervical nerves: The neurapraxia (i.e., injury to a nerve resulting in paralysis that recovers completely in a short period of time, known as “stingers” or “burners”) that occurs in contact sports is thought to result from injury to the cervical nerve roots (usually thought to be C5 or C6) or the brachial plexus. These injuries are quite common in American football with 65% of college players experiencing a stinger/burner during their 4 years of football (Safran, 2004). Narrowing of the vertebral canal (spinal stenosis) of the cervical region (defined as a Torg ratio <0.8) results in a threefold increase in the risk of stingers from an extension-compression mechanism in football players (Meyer et al., 1994). The Torg ratio is the anterior-to-posterior distance between the vertebral body and the spinous process–laminar junction at the narrowest region of the cervical vertebral canal divided by the anterior-to-posterior dimension of the vertebral body at the same level (Torg et al., 1986). The measurements are made from lateral extension x-rays. The presence of stenosis also increases the incidence of a complicated recovery from these injuries (Meyer et al., 1994).
Spinous Process
The spinous process of a typical cervical vertebra is short and bifid posteriorly. It is bifid because it develops from two separate secondary centers of ossification. This morphology is unique to cervical spinous processes. The “terminal tubercles” of the bifid spinous process are frequently of unequal size and allow for attachment of the ligamentum nuchae (Standring et al., 2008) and many of the deep extensor muscles of the spine (semispinalis thoracis and cervicis, multifidi cervicis, spinalis cervicis, and interspinalis cervicis muscles).
Cervical spinous processes, as with spinous processes throughout the spine, may deviate from the midline, making the determination of structural defects, fractures, and dislocations more challenging (Standring et al., 2008). The length of the spinous processes decreases from C2 to C4 and then increases from C4 to C7 (Panjabi et al., 1991).
Intervertebral Foramina
The left and right intervertebral foramina (IVFs) in the cervical region lie between the inferior and superior vertebral notches of adjacent cervical vertebrae (see Fig. 5-3, C and D). They face obliquely anteriorly at approximately a 45-degree angle from the midsagittal plane for the upper and middle cervical IVFs and approximately a 55-degree angle to the midsagittal plane for the lower cervical IVFs (Marcelis et al., 1993). The IVFs also are directed inferiorly at approximately a 10-degree angle to a horizontal plane passing through the superior vertebral end plate. The specific borders and contents of the IVF are discussed in Chapter 2. However, the uncinate processes, which help to form the anterior border of the IVFs, are unique to the cervical region. The cervical IVFs, as with those of the thoracic and lumbar regions, can best be considered as neural canals because they are 4 to 6 mm in length. They are almost oval in shape. The average height of the cervical IVFs is 8.1 mm and the average width is 5.6 mm. Foraminal height increases from C3-4 (7.4 ± 1.1 mm) to C7-T1 (8.6 ± 1.0 mm), as does width (C3-4 = 4.5 ± 0.8 mm, C7-T1 = 7.0 ± 1.1 mm). The dimensions of C2-3 left and right IVFs are more similar to those of the middle to lower cervical IVFs (height = 8.1 ± 1.2 mm, width = 5.2 ± 0.9 mm) (Ebraheim, Biyani, & Salpietro, 1996).
The IVFs change dimensions with movement. Flexion of 20 degrees, 30 degrees, and full flexion from the neutral position increase the superior to inferior IVF diameter by 8%, 10%, and 31%, respectively; and extension of 20 degrees, 30 degrees, and full extension from the neutral position decreases IVF diameter by 10%, 13%, and 20%, respectively (Yoo et al., 1992; Muhle et al., 2001). Therapeutic traction also increases the IVF area and height (18.9% and 10.4% increases, respectively, with 15-kg traction weight) (Liu et al., 2008). The pressure within the IVF also increases during extension, and elevation (abduction) of the upper extremity decreases pressure within the IVF (Farmer & Wisneski, 1994). Relief of pain radiating distally in the upper extremity after abduction of the arm is known as a positive Bakody’s sign, and the decreased IVF pressures associated with abduction of the upper extremity may explain the mechanism of this sign. Ipsilateral axial rotation of 20 and 40 degrees results in foraminal narrowing of up to 15% and 23%, respectively; and contralateral axial rotation of 20 and 40 degrees results in foraminal widening of up to 9% and 20%, respectively (Muhle et al., 2001).
The dorsal and ventral roots (medially) or the spinal nerve (laterally) and the dural root sleeve account for 35% to 50% of the cross-sectional area of a typical cervical IVF (Sunderland, 1974). When the cervical spine is in the neutral position, the dorsal and ventral roots are located in the inferior portion of the IVF at or below the disc level (Pech et al., 1985). Epidural fat and blood vessels are found in the superior aspect of the IVF. The dorsal root and dorsal root ganglion are located posterior to and slightly above the ventral root. The dorsal root is also in contact with the superior articular process. The dorsal root ganglion is associated with a small notch on the anterior surface of the superior articular process in the cervical region. The ventral root contacts the uncinate process, and the dorsal and ventral roots are separated from each other by adipose tissue (Pech et al., 1985). This adipose-filled region between the dorsal and ventral roots has been called the interradicular foramen or cleft and can be seen on MRI (Yenerich & Haughton, 1986).
Stenosis of the intervertebral foramina: Narrowing (stenosis) of the IVFs can occur for a variety of reasons. Such stenosis can result in compression of the dorsal and ventral rootlets or roots, or dorsal root ganglion. This can lead to decreased muscle strength in the muscles innervated by the affected ventral roots, or pain radiating along the distribution of the nerves composed of the peripheral processes of the neurons found in the dorsal roots. Pain that results from compression of the dorsal rootlets, roots, or the dorsal root ganglion is known as radicular pain (see Chapter 11). Recall that the rootlets and roots are surrounded by the meninges and, more specifically, the dorsal and ventral roots and dorsal root ganglion are surrounded by a dural root sleeve. As mentioned, the cervical dural root sleeves are infundibular (funnel shaped), being widest proximally, and compression of the neural elements usually occurs within the proximal IVF. The structures that most frequently cause stenosis and compression of the neural elements from the anterior are the IVDs, and bone spurs (osteophytes) from the uncinate processes or vertebral bodies. Structures that most frequently cause foraminal stenosis and compress the neural elements from the posterior are fibrous tissue in the IVF, ligamentum flavum that has thickened or buckled, and osteophytes from the superior or inferior articular process (Bailey & Casamajor, 1911; Hadley, 1957; Bland, 1989; Humphreys et al., 1998; Tanaka et al., 2000). Other possible causes of cervical IVF stenosis and radicular pain include fibrous adhesions within the IVF and inflammation of the nerve roots caused by the adhesions, edema, and congestion of the blood vessels within the cervical IVFs (Hadley, 1957). Radicular pain of the cervical dorsal roots is most common in the C6, C7, and C8 roots, with C6 accounting for 26%, C7 accounting for 61%, and C8 accounting for 8% of radicular pain of cervical origin (Murphy, Simmons, & Brunson, 1973). One reason for the lower incidence of radiculopathy from the C8 nerve roots is that they lie above the C7-T1 IVDs, whereas the C6 and C7 nerve roots are located directly posterior to the C5-6 and C6-7 IVDs, respectively (Tanaka et al., 2000).
Stenosis of the IVFs can be evaluated with standard x-rays, CT scanning, or MRI. Direct oblique MRI provides more accurate assessment of all of the borders of the cervical IVFs than standard sagittal images (Cramer et al., 2002). Kinalski and Kostro (1971) found the area of the cervical IVFs (as recorded from plain oblique radiographs) to correlate with age and patient symptoms. Individuals 20 to 40 years of age had larger IVFs than those older than 40. Also, a smaller IVF size was found among patients with chronic neck pain. These findings were confirmed by Humphreys and colleagues (1998), who found that the left-to-right width of the IVFs decreased with age and that this dimension also was smaller in individuals with chronic neck pain with radiculopathy.
Enlargement of the intervertebral foramina: An IVF may enlarge as a result of various pathologic conditions. The most common cause of significant pathologic enlargement of the IVF is the presence of a neurofibroma. Less frequently enlargement may be caused by meningioma, fibroma, lipoma, herniated meningocele, a tortuous vertebral artery (Danziger & Bloch, 1975), congenital absence of the pedicle with malformation of the TP (Schimmel, Newton, & Mani, 1976), and chordoma (Wang et al., 1984).
External Aspect of the Occipital Bone
The external surface of the occipital bone is so intimately related to the spine (direct articulation and ligamentous attachments with the atlas and ligamentous attachments with the axis) that it is included in this section on the cervical region.
The external aspect of the occipital bone consists of three different regions: squamous, left and right lateral, and basilar. These three regions are discussed separately.
Squamous Part
The squamous part of the occipital bone (occipital squama) is located posterior to the foramen magnum (Fig. 5-8). The part of the occipital bone that forms the most posterior aspect of the foramen magnum is known as the opisthion. The most prominent feature of the occipital squama is the external occipital protuberance (EOP). This mound, whose summit is known as the inion, serves as the attachment site for the medial insertion of the trapezius muscle. The external occipital crest extends inferiorly from the EOP.
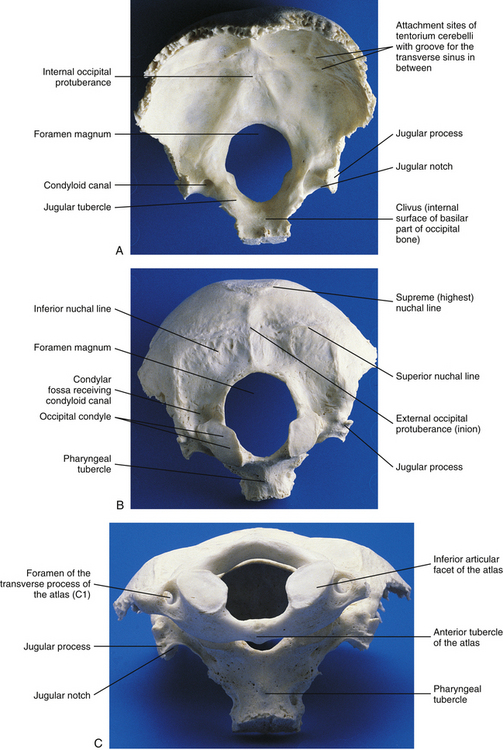
FIG. 5-8 A, Superior, or internal, and, B, inferior, or external, views of the occiput. C, Atlas articulating with the occiput.
The squamous part of the occipital bone also has several markings formed by muscular and ligamentous attachments. Extending laterally from the EOP are two pairs of nuchal lines. The first is present only occasionally and is known as the highest (supreme) nuchal line. The second is almost always present and is known as the superior nuchal line. The highest nuchal line, when present, extends superiorly and laterally from the EOP. It is formed by attachment of the occipital belly of the occipitofrontalis (epicranius) muscle. The superior nuchal line extends almost directly laterally from the EOP and is formed by the attachment of the trapezius and sternocleidomastoid (SCM) muscles. A third nuchal line called the inferior nuchal line extends laterally from the external occipital crest approximately midway between the EOP and foramen magnum. Several muscles attach above and below the inferior nuchal line (Table 5-2).The posterior atlanto-occipital membrane attaches to the most inferior aspect of the occipital squama, which is the posterior border of the foramen magnum.
Table 5-2
| Region | Muscles and Ligaments Attached |
| Squamous | Trapezius Sternocleidomastoid Occipital belly of occipitofrontalis Splenius capitis Semispinalis capitis Obliquus capitis superior Rectus capitis posterior major and minor Posterior atlanto-occipital membrane |
| Lateral | Rectus capitis lateralis Alar ligament |
| Basilar | Rectus capitis anterior Longus capitis Superior pharyngeal constrictor Anterior atlanto-occipital membrane Apical ligament of odontoid process Superior (upper) band of cruciform ligament Tectorial membrane |
Lateral Parts
The left and right lateral portions of the occipital bone are located to the sides of the foramen magnum. They include the left and right occipital condyles, jugular processes, and jugular notches.
The two occipital condyles are convex structures located on each side of the foramen magnum. Each condyle follows the contour of the large foramen and protrudes inferiorly, anteriorly, and medially (see Fig. 5-8). Each possesses a hyaline cartilage–lined articular facet. The left and right occipital condyles fit snugly into the superior articular facets of the atlas, and the left and right atlanto-occipital articulations allow for flexion, extension, and lateral flexion of the occiput on the atlas. The facetal surface of occipital condyles may be constricted in the center and occasionally may be completely divided (bipartite). When this occurs, the superior articular facet of the atlas matches the articular facet of the corresponding occipital condyle and also is constricted or bipartite (Gottleib, 1994).
The jugular notch is a groove along the lateral margin of each side of the occiput. This groove helps to form the large jugular foramen of the same side by lying in register with the jugular fossa of the temporal bone.
The jugular notch is bounded laterally by the jugular process. The jugular process is an anterior projection on the lateral aspect of each side of the occiput. Each one helps to form the posterolateral margin of the jugular foramen of the same side. The rectus capitis lateralis muscle, which helps to laterally flex the occiput on the atlas, attaches to this process.
Basilar Part
The basilar region of the occipital bone extends anteriorly from the foramen magnum. The part of the occipital bone that forms the most anterior aspect of the foramen magnum is known as the basion. The basilar region of the occipital bone meets the basilar portion of the sphenoid bone, and together the internal surface of the two basilar processes is known as the clivus.
The superior constrictor muscle of the pharynx attaches to the distinct pharyngeal tubercle, which is located in the center of the external surface of the basiocciput. The rectus capitis anterior muscle attaches just in front of the occipital condyle, and the longus capitis muscle attaches anteriorly and laterally to the pharyngeal tubercle. The anterior atlanto-occipital membrane attaches just in front of the foramen magnum. The apical ligament of the odontoid process attaches to the rim of the foramen magnum, and the superior (upper) band of the cruciform ligament attaches to the surface of the clivus, covered posteriorly by the clival attachment of the tectorial membrane. The transition of the spinal dura to the meningeal layer of the cranial dura occurs just posterior to the tectorial membrane.
Congenital Anomalies of the Occiput and Clinical Considerations
The craniovertebral region is associated with many variants and anomalies. These variations in normal anatomy and related anomalies are thought to primarily result from genetic transmission, although environmental factors may influence their development (Taitz, 2000).
Normally the occipital sclerotomes and the first cervical sclerotome are incorporated into the occiput (see Chapter 12). If these sclerotomes are incompletely incorporated, then remnants of an occipital vertebra may develop. These remnants can result in a variety of structures in the fully developed skull, including a paracondylar process, epitransverse process, hypocondylar arch, or third occipital condyle (Fig. 5-9).
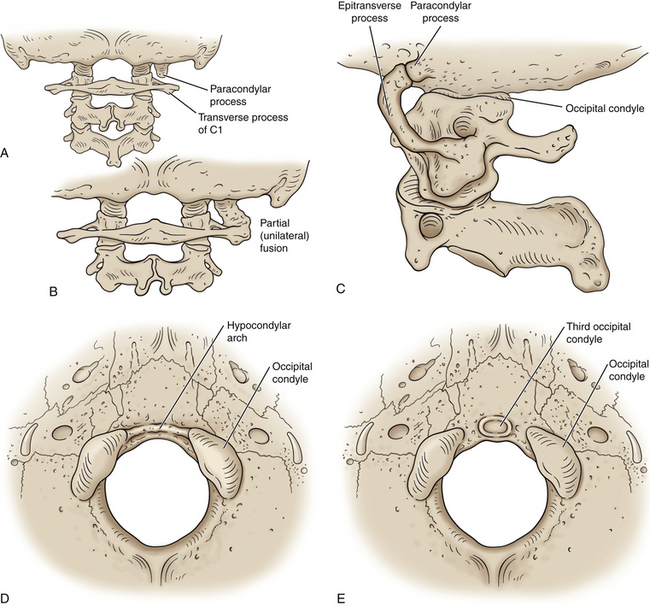
FIG. 5-9 Atlanto-occipital anomalies. A, Posterior view of a paracondylar process seen lateral to the right occipital condyle. B, Paracondylar process extending to the atlas (C1) on the right side with unilateral (i.e., partial) fusion of the atlas to the occiput. C, Lateral view showing an epitransverse process that extends anteriorly and superiorly from the transverse process of the atlas to form an articulation with a paracondylar process of the occiput. D, Inferior view of the occiput showing a hypocondylar arch that extends between the anterior and medial aspects of the left and right occipital condyles. E, When the hypocondular arch does not reach the occipital condyles the term “third occipital condyle” is used to describe the anomaly. The odontoid process or anterior arch of the atlas will sometimes, but not always, articulate with a third occipital condyle.
A paracondylar process is a bony process that protrudes inferiorly from the occiput just lateral to an occipital condyle (Fig. 5-9, A and B). This type of process can cause decreased or aberrant motion of the upper cervical region and has been listed as a cause of cervical torticollis of skeletal origin. An epitransverse process is a bony projection from the transverse process of the atlas that extends superiorly toward the occiput. In some cases an epitransverse process can form a crude joint (pseudarthrosis) with a paracondylar process descending from the occiput (Fig. 5-9, C).
A hypocondylar arch is a prominent ridge coursing along the anterior margin of the foramen magnum (Fig. 5-9, D). If the ridge is incomplete, the term “third occipital condyle” is used to describe the elevation along the anterior margin of the foramen magnum (Fig. 5-9, E). The third occipital condyle can form a pseudarthrosis with the odontoid process or anterior arch of the atlas (Taitz, 2000; von Ludinghausen et al., 2005). In addition, accessory ossicles are occasionally found between the anterior arch of the atlas (or the odontoid process) and the occiput. Rarely an accessory ossicle forms between the posterior arch of the atlas and the occiput posterior to the foramen magnum. Such an accessory posterior ossicle is called an os, or processus, Kerckring (von Ludinghausen et al., 2005).
Other common anomalies of the craniovertebral region are associated with a lack of segmentation of the occipital and upper cervical somites, and include various degrees of assimilation, or fusion, of the atlas with the occiput. Such assimilations can be almost complete fusions, but usually appear as asymmetric fusion of one lateral mass to the adjacent occipital condyle. However, both lateral masses and the anterior arch of C1 may fuse with the occiput, but the posterior arch usually is spared from such fusion, allowing room for the vertebral artery to enter the foramen magnum.
The various forms of fusion of the atlas with the occiput frequently are accompanied by ascension of the odontoid process either into the foramen magnum or to just the inferior border of the foramen magnum. These locations of the odontoid process can result in partial compression of the inferior aspect of the medulla or the first cervical spinal cord segment (Taitz, 2000).
The various forms of atlantal fusions also can be accompanied by superior deviation of the inferior aspect of the occiput, known as basilar invagination, or basilar impression. Basilar invagination can result in neurologic signs and symptoms of cerebellar and lower cranial nerve dysfunction. In fact, anomalies of the occipitocervical region in general are not infrequently associated with neurologic disorders related to the region of the medulla and upper cervical spinal cord segments. These disorders usually manifest themselves later in life (after 30 years of age), unless they are triggered by trauma to the upper cervical region in earlier years (Taitz, 2000).
Because the somites (see Chapter 12), branchial arches, and mesonephros are developing at nearly the same time, anomalies of the craniovertebral region frequently are accompanied by additional anomalies of the face, ear, and other cervical vertebrae. Less frequently, anomalies of the craniovertebral region are accompanied by anomalies of the thoracic and lumbar vertebrae (e.g., supernumerary thoracic vertebra and transitional lower lumbar segments) and with congenital disorders of the urinary system (Taitz, 2000).
Atypical and Unique Cervical Vertebrae
The atypical cervical vertebrae are C1 and C2. C7 is unique. C6 is considered to have unique characteristics but remains typical. The distinctive features of these vertebrae are discussed in the following sections.
Atlas (First Cervical Vertebra)
The most superior atypical vertebra of the spine is the first cervical vertebra (Fig. 5-10). Given the name atlas, after the Greek god, this vertebra actually does function to support a round sphere (the head). The fully developed atlas comprises two arches (anterior and posterior) separated by two laterally placed pieces of bone known as the lateral masses. The lateral masses, in turn, have a TP projecting from their sides. The atlas develops from three primary centers of ossification, one in each lateral mass and one in the anterior arch (see Fig. 13-9). The centers located in the lateral masses are the first to appear, and are formed by approximately the seventh week after conception. These centers develop posteriorly into the future posterior arch of the atlas, where they usually unite by approximately the fourth year of life. Occasionally these ossification centers fail to unite posteriorly, leaving a cartilaginous bridge. This cartilaginous bridge is radiolucent on standard x-ray films. Such radiolucency must be differentiated from a fracture in trauma patients.
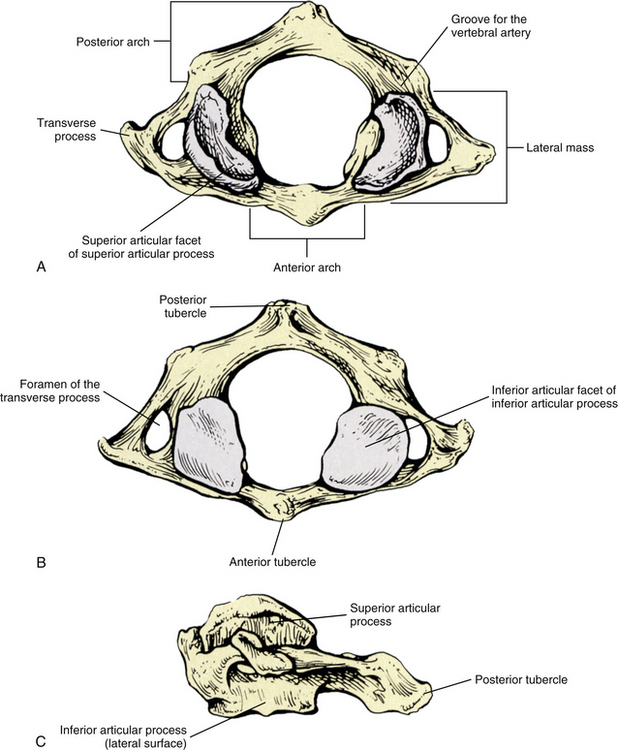
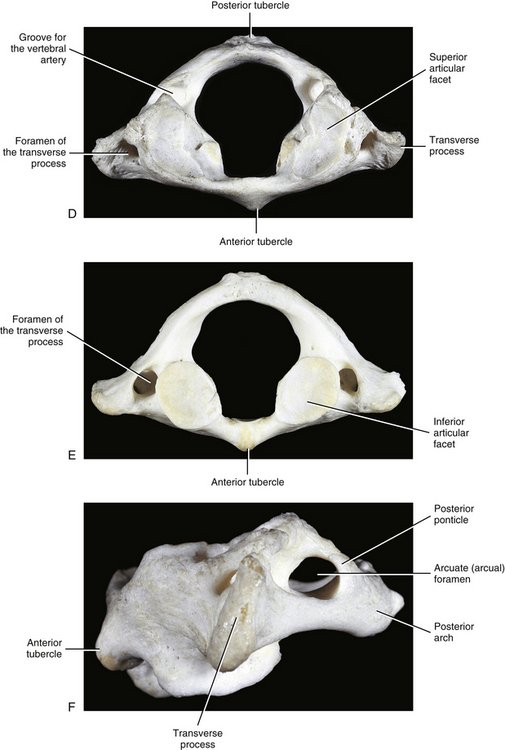
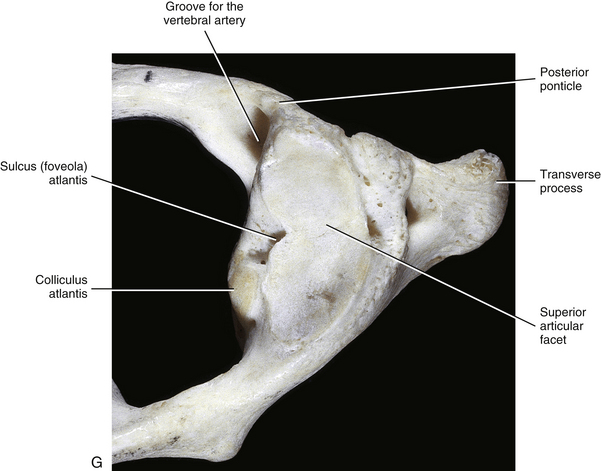
FIG. 5-10 A, Superior, B, inferior, and, C, lateral views of the first cervical vertebra—the atlas. D, Superior, E, inferior, and, F, lateral views of the first cervical vertebra—the atlas. G, Close-up (superior view) of the lateral mass of the atlas. Notice the colliculus atlantis and sulcus (foveola) on the medial surface of the lateral mass.
Anterior Arch
The anterior arch of the atlas develops from a bridge of tissue that connects the two lateral masses of the atlas in the embryo. This bridge is known as the hypochordal arch, and although the hypochordal arch is found throughout the spine embryologically, the anterior arch of the atlas is the only place where it persists into adulthood. The primary ossification center of the anterior arch usually appears by the end of the first year of postnatal life and fuses with the left and right lateral masses between the ages of 7 and 9 years (Fesmire & Luten, 1989).
The anterior arch is the smaller of the two atlantal arches (see Fig. 5-10). It possesses an elevation on its anterior surface known as the anterior tubercle. This tubercle serves as the attachment site for the anterior longitudinal ligament centrally and the superior oblique fibers of the longus colli muscle slightly laterally.
The posterior surface of the anterior arch (see Fig. 5-10) contains a smooth articulating surface known as the facet for the dens (odontoid). This facet is covered with hyaline cartilage and articulates with the anterior surface of the odontoid process as a diarthrodial joint. Because the atlas has no vertebral body, the odontoid process of the axis occupies the region homologous to the body of the atlas. Consequently, the atlas is oval in shape and can pivot easily around the odontoid process at the diarthrodial joint between this process and the anterior arch of C1.
Posterior Arch
The posterior arch is larger than the anterior arch and forms approximately two thirds of the ring of the atlas. The larger posterior arch contains an elevation on its posterior surface known as the posterior tubercle. This tubercle may be palpated in some individuals. It serves centrally as an attachment site for the ligamentum nuchae (Standring et al., 2008) and also as the origin for the rectus capitis posterior minor muscle.
The first left and right ligamenta flava attach to the lower border of the posterior arch of the atlas. The ligamenta flava are discussed in more detail later in this chapter. The lateral aspects of the superior surface of the posterior arch are extremely thin and “dug out.” These dug-out regions are known as the left and right grooves for the vertebral arteries. Each groove allows passage of the vertebral artery, vertebral veins, and suboccipital nerve of the same side. The suboccipital nerve is the dorsal ramus of C1 and is located between the vertebral artery and posterior arch. The groove for the vertebral artery has been found to be covered by an arch of bone in approximately 32% to 37% of subjects studied (Taitz & Nathan, 1986). This results in the formation of a foramen, sometimes called the arcuate or arcual foramen (Fig. 5-10, F). Each vertebral artery reaches its respective groove as it courses around the superior articular process and comes to lie on top of the posterior arch. The posterior atlanto-occipital membrane attaches to each side of the groove, and it is the lateral and inferior edge of this membrane that may ossify to create an arcuate foramen (see Fig. 5-16). When ossification occurs, the bone bridge (partial or complete) that is created is known as a posterior ponticle (Fig. 5-10, F). A study of 672 atlas vertebrae found that 25.9% had a partial posterior ponticle and 7.9% had a complete posterior ponticle (Taitz & Nathan, 1986). A higher incidence of complete posterior ponticles (28.4%) has been reported in the Kenyan population (Karau et al., 2010), whereas the incidence of partial or complete posterior ponticles in the Korean population appears to be lower (15.6%; males 19.3%, females 12.8%) (Hong et al., 2008). Taitz and Nathan (1986) reported a much higher incidence (57%) of partial or complete ponticle formation in a Middle Eastern population, possibly because this population customarily carried heavy loads on their heads. However, these authors stated that further study is necessary to determine with certainty the cause of the high incidence of posterior ponticles in this population.
The incidence of complete versus partial posterior ponticles is higher with increased age, indicating that posterior ponticles grow in a similar manner to other bone spurs of the vertebral column (e.g., osteophytes), that is, being more common with increased age (Hong et al., 2008).
A significant relationship has been found between the presence of a posterior ponticle and migraine headache without aura. However, the reason for this relationship is unclear. Speculation includes ischemic compression of the vertebral artery or tension on the dura mater in the region of the occiput and atlas (Wight, Osborne, & Breen, 1999).
Occasionally a bone bridge for the vertebral artery develops laterally between the superior articular process of the atlas and the TP. Such a process is known as a lateral ponticle (ponticulus lateralis) (Buna et al., 1984). Lateral ponticles are found in approximately 4% of atlas vertebrae and are more common on the right side. They are also more common if a posterior ponticle is also present (13% if a posterior ponticle is present) (Taitz & Nathan, 1986; Karau et al., 2010). Regardless of whether the ponticle is posteriorly or laterally placed, they are usually more than 12 mm in length and thicker than 1 mm. Some controversy surrounds whether posterior and lateral ponticles are congenital or are a part of the aging process. Taitz and Nathan (1986) found that partial ponticles were predominant in the specimens of 10- to 30-year-old individuals, and complete posterior ponticles usually were found in specimens from 30- to 80-year-old individuals. This indicates that at least posterior ponticles are created by ossification of the lateral-most portion of the posterior atlanto-occipital membrane as some people age. Even though these ponticles also have been implicated in some cases of vertebrobasilar arterial insufficiency (Buna et al., 1984), their clinical significance remains a matter of debate.
Lateral Masses
Located between the anterior and posterior arches are the left and right lateral masses. Each mass consists of a superior articular process, an inferior articular process, and a transverse process. The average superior to inferior height of a lateral mass is 14.09 mm (±1.92 mm) (Dong et al., 2003). Each superior articular process (SAP) is oriented so that the anterior aspect is more medially positioned than the posterior aspect. Recall that the anterior aspect of each lateral mass serves as the origin for the rectus capitis anterior muscle. The medial surface of each lateral mass has a small tubercle for attachment of the transverse atlantal ligament. This tubercle has been called the colliculus atlantis (see Fig. 5-10, G), and usually develops by the age of 13 years. A prominent depression that has been called the sulcus atlantis (foveola atlantis) is found immediately posterior to the colliculus. A nutrient artery to the lateral mass that branches from the third part of the vertebral artery usually enters the lateral mass through a nutrient foramen in the sulcus atlantis. The sulcus atlantis can be seen on CT scans taken in the horizontal (axial) plane and the colliculus atlantis can be seen on both axial CT scans and standard anteroposterior open mouth radiographs of the atlas (Weiglein & Schmidberger, 1998).
The SAP of each lateral mass has an average maximum left-to-right width of 15.47 mm (±1.19 mm) and an average anterior-to-posterior dimension of 17.21 mm (±0.93 mm) (Dong et al., 2003). The SAPs are irregular in shape. In fact, the hyaline-lined superior articular facet of each SAP usually has the appearance of a peanut. That is, it is narrow centrally and wider anteriorly and posteriorly. The SAP is quite concave superiorly and faces slightly medially to accommodate the convex occipital condyle of the corresponding side. The concavity of the SAP increases with age (Hallgren, Cattrysse, & Zrull, 2011). The joint between the occiput and atlas is categorized as a condyloid, diarthrodial joint, although some authors describe it as being ellipsoidal in type because of its shape (Standring et al., 2008). The primary motion at this joint is anteroposterior rocking (flexion and extension). In addition, a small amount of lateral flexion occurs at this articulation.
Asymmetry of the left and right SAPs of the atlas is the rule rather than the exception. Approximately 63% of SAPs of atlases are “grossly asymmetric,” and approximately 11% are slightly asymmetric. Approximately 37% of grossly asymmetric SAPs have three or four distinct articular facets (two on one side and one on the other, or two on each side). In addition, the depth of the concavity of the SAPs is quite variable, ranging from being almost flat (<3-mm maximum depression) to being very concave (≥5-mm maximum depression) (Gottleib, 1994). Further investigation is necessary to determine the effects of asymmetry on motion between the occiput and atlas.
The inferior articular process of each lateral mass of the atlas presents as a regularly shaped oval. In fact, it is almost circular in many cases. This process is flat or slightly concave (Standring et al., 2008) and faces slightly medially. Hyaline cartilage lines the slightly smaller inferior articular facet of the articular process. The average maximum left-to-right width of the inferior articular facet is 17.90 mm (±1.18 mm) (Dong et al., 2003). The inferior articular facet articulates with the superior articular facet of C2. A loose articular capsule attaches to the rim of the corresponding articular facets, surrounding the lateral C1-2 joint. This loose capsule allows approximately 45 degrees of unilateral rotation to occur at each atlanto-axial joint. This joint is categorized as a planar diarthrodial articulation (typical joint type for Z joints).
Transverse Processes
The left and right TPs of the atlas are large and may be palpated between the mastoid process and the angle of the mandible (see Chapter 1). Each projects laterally from the lateral mass and acts as a lever by which the muscles that attach to it may rotate the head. Because of the large size of the TPs, the atlas is wider than all the cervical vertebrae except C7. The width ranges from approximately 65 to 76 mm in females and from 74 to 95 mm in males (Standring et al., 2008). Although the size and shape of the left and right transverse processes frequently are asymmetric (Pei-Feng et al., 2011), each is usually composed of only a single lateral process (rather than having anterior and posterior tubercles, as is the case with typical cervical vertebrae). The single lateral process of the C1 TP is almost completely homologous to the posterior roots, or bars, of the other cervical vertebrae. In fact, the TP of the atlas can be considered to be composed of a posterior root and a small portion of the intertubercular lamella (Standring et al., 2008). A foramen for the vertebral artery, which also provides passage for the vertebral veins and the vertebral artery sympathetic nerve plexus, also is found within each transverse process. This foramen of the TP of C1 is the largest of the cervical spine (Taitz, Nathan, & Arensburg, 1978). The anterior aspect of the C1 foramen of the TP fails to close (i.e., remains open) in 10.2% of individuals (bilateral occurrence 45.1%, unilateral 54.9%). There is no known clinical significance to this anatomic variation (Billmann & Le Minor, 2009).
In addition to its relationship with the vertebral vessels, each TP of the atlas is also the site of muscle attachments. These muscles include the rectus capitis lateralis, obliquus capitis superior, obliquus capitis inferior, levator scapulae, splenius cervicis, and scalenus medius muscles. Each TP also is related to the C1 spinal nerve. Although the dorsal ramus of this spinal nerve (the suboccipital nerve) provides motor innervation to the suboccipital muscles, the ventral ramus passes laterally around the lateral mass, remaining medial to the vertebral artery and the rectus capitis lateralis muscle. It courses between the rectus capitis lateralis and anterior muscles and then descends anterior to the atlas and is joined by the ascending branch of the ventral ramus of C2. The nerves of the cervical region are discussed in more detail later in this chapter.
Vertebral Foramen
The large vertebral foramen of C1 usually has a greater anteroposterior than transverse diameter (Le Minor, Kahn, & Di Paola, 1989). The anteroposterior dimensions of the C1 vertebral foramen can be divided into thirds (Fig. 5-11), with one third filled with the odontoid process of C2, one third filled with the spinal cord, and one third being “free space.” The free space actually is filled with epidural adipose tissue, vessels, ligaments, meninges, and the subarachnoid space. This division of the vertebral foramen of the atlas into three parts is sometimes known as Steel’s rule of thirds (Foreman & Croft, 1992). Rarely the vertebral foramen of C1 can be narrowed by a hypoplastic posterior arch. This narrowed C1 vertebral foramen can result in compression of the spinal cord (myelopathy) (Musha & Mizutani, 2009).
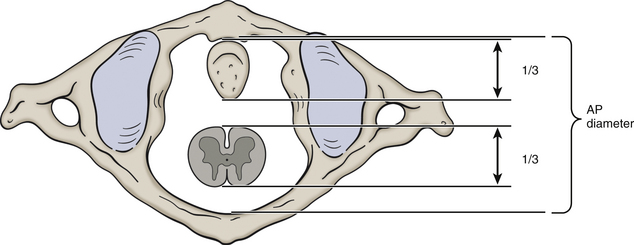
FIG. 5-11 Steel’s rule of thirds.The C1 vertebral foramen can be divided into thirds (“Steel’s rule of thirds”) where one third of the foramen is filled with the odontoid process, one third with the spinal cord, and the final third is referred to as “free space.” The “free space” is actually filled with the epidural venous plexus and surrounding adipose tissue (i.e., epidural fat); dura, arachnoid and cerebrospinal fluid in the subarachnoid space; and upper cervical ligaments (tectorial membrane and transverse portion of the cruciform ligament).
Axis (Second Cervical Vertebra)
The second cervical vertebra, the axis or epistropheus, also is atypical. This vertebra develops from five primary and five secondary centers of ossification. The primary centers are distributed as follows: one in the vertebral body, two in the neural arch (one on each side), and two in the dens (odontoid process, see the following section). One secondary center of ossification is associated with the odontoid process, another is associated with the inferior aspect of the vertebral body, two are associated with the transverse processes (one for each, left and right), and one is located at the posterior tip of the spinous process.
The major distinguishing features of the axis are the prominent dens (odontoid process), superior articular processes, and transverse processes (Fig. 5-12). These distinguishing features are discussed in the following sections. In addition, the vertebral foramen of C2 is large.
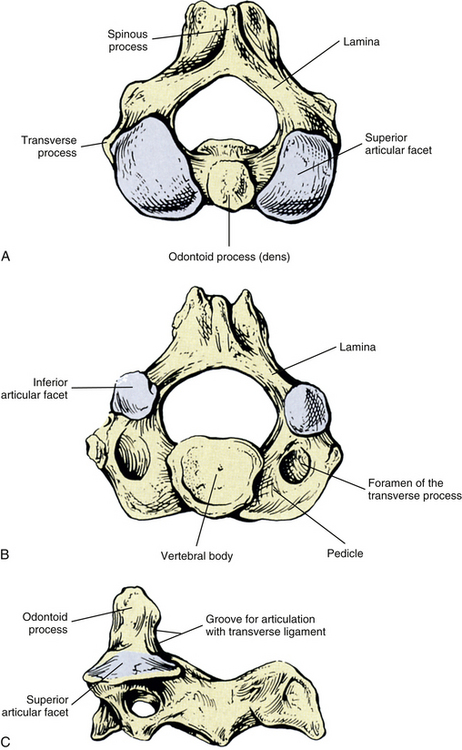
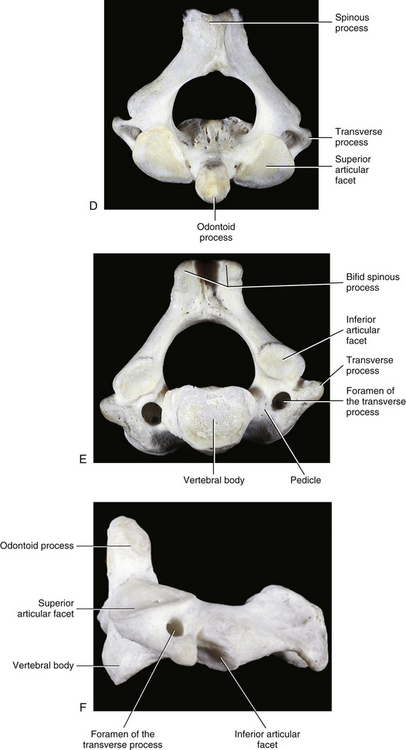
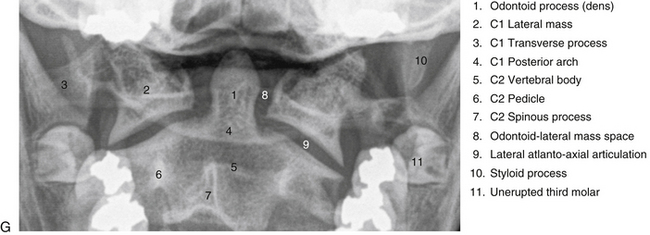
FIG. 5-12 A, Superior, B, inferior, and, C, lateral views of the second cervical vertebra—the axis. D, Superior, E, inferior, and, F, lateral views of the second cervical vertebra—the axis. G, Anterior-posterior “open mouth” (APOM) view of the upper cervical region showing the atlas, axis, and related bony structures. (G, Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
Dens (Odontoid Process)
Also known as the odontoid process, the dens develops from two laterally placed primary centers of ossification and an apical secondary center of ossification (see Fig. 13-10). The two primary centers appear in utero and usually fuse in the midline by the eighth fetal month (Fesmire & Luten, 1989). The united primary ossification centers then normally fuse along their inferior outer rim to the vertebral body of C2 by approximately age 3 to 6 years. The fusion line between the odontoid and the body of C2 usually is visible on x-ray film until approximately age 11, and one third of individuals retain the line of fusion throughout life. This line is frequently confused with a fracture (Fesmire & Luten, 1989). Inside the rim of attachment between the dens and the body of C2 a small disc is present, which persists until late in life. This disc frequently can be seen on sagittal MRI scans. This area of fusion between the odontoid and the body of C2 is known as the subdental synchondrosis.
The apical secondary center of ossification first appears at 3 to 6 years of age. It is V or cuneiform in shape, forming a deep cleft between the primary centers of ossification (see Fig. 13-10, D). This secondary center unites with the remainder of the odontoid process usually by age 12. When seen on x-ray film before or in the absence of fusion, the small bone fragment is known as a persistent ossiculum terminale (or simply ossiculum terminale, if before age 12) and also can be difficult to distinguish from a fracture.
The fully developed odontoid process (dens) is peg shaped with a curved superior surface. In approximately 14% to 26% of the population it deviates slightly to the left or right sides (Pfirrmann et al., 2001; Ji-Hong et al., 2010). The height of the odontoid process is 15.5 mm (±1.8 mm) in males and 14.6 mm (±1.5 mm) in females. Its anterior-to-posterior diameter is 10.3 mm (±0.7 mm) in males and 9.6 mm (±0.9 mm) in females (Xu et al., 1995). The dens has a hyaline-lined articular facet on its anterior surface. This facet articulates with the corresponding facet on the posterior surface of the anterior arch of the atlas. The posterior surface of the dens has a groove at its base formed by the transverse atlantal ligament (transverse portion of the cruciform ligament). The transverse ligament forms a synovial joint with the groove on the posterior surface of the dens. Together the complex of anterior and posterior joints between the atlas, odontoid, and transverse ligament is classified as a trochoid (pivot) diarthrodial (synovial) joint. This joint allows the atlas to rotate on the axis through approximately 45 degrees of motion in each direction (left and right). The sides of the odontoid process above the groove formed by the transverse ligament are flat and serve as attachment sites for the left and right alar ligaments. The apical odontoid ligament attaches to the top of the odontoid process. The ligaments and joints of the cervical spine are discussed later in this chapter.
The odontoid process receives a rich arterial supply. Before entering the foramen of the transverse process of their respective side, both the left and right vertebral arteries give an anterior and posterior ascending artery of the axis that ascend the anterior and posterior surfaces of the odontoid process, respectively. The four ascending arteries (left and right anterior and posterior) unite superiorly to form an apical arterial arcade of the odontoid process. In addition, before entering the carotid canal, the superior aspect of the extracranial component of the internal carotid artery sends branches to the anterior surface of the odontoid process and additional branches that reinforce the portion of the anterior ascending artery of the axis that supplies the odontoid process (Schiff & Parke, 1973). The remnant of an intervertebral disc (subdental synchondrosis) between the odontoid process and the body of C2 does not allow a meaningful vascular exchange between the two regions, which explains the slow healing of many type II odontoid fractures and the persistence of os odontoideum (see following sections) (Menezes & Traynelis, 2008).
Os odontoideum: Rarely the odontoid is not properly fused with the body of C2 or is united by only a rim of cartilage. Therefore its appearance on x-ray film is that of a free and unattached odontoid (see Figure 13-12). This unfused odontoid is known as an os odontoideum (OO). The precise incidence of OO is unknown, although it is considered to be a rather rare phenomenon with the exception of Down syndrome children in whom the incidence is approximately 6% (Hankinson & Anderson, 2010). Previously OO was thought to result from a lack of fusion of the odontoid process with the body of the axis during early childhood development; however, this condition is now considered usually to result from a fracture of the base of the odontoid (type II odontoid fracture) during the early postnatal period (Lefebvre et al., 1993; Lohiya, 1993; Verska & Anderson, 1997). The natural history of OO is unknown, and OO can exist without symptoms. However, relatively minor trauma can cause a previously pain-free OO to become symptomatic (Lohiya, 1993; Sosner, Fast, & Kahan, 1996). In some cases, OO can lead to severe neurologic deficits. However, limited neck motion and neck pain in the upper cervical region are the most common signs and symptoms of an unstable OO, and less often torticollis (neck laterally flexed and rotated to the same side) and headache may be present as well. Radiographic examination is an important part of the accurate diagnosis of an unstable OO. Although OO appears as a translucent line at the base of the dens on standard x-rays, such a translucent line seen in the x-rays of children corresponds to the normal appearance of the cartilaginous attachment (growth plate) of the dens to the body of C2. This growth plate closes in 50% of children by the age of 4 years and should be closed in all children by 6 years of age, although the translucent line frequently persists until the age of 11 years. Therefore further investigation with lateral projection and flexion-extension x-rays is warranted in both adults and children suspected of having an OO, to reveal whether or not the odontoid is unstable. The space between the anterior aspect of the dens and anterior arch of the atlas (atlanto-odontal interspace) is normal in an unstable OO. However, the distance between lines drawn along the posterior aspect of the anterior arch of the atlas and anterior aspect of the body (not the odontoid) of C2 increases in flexion as compared with the neutral position in an unstable OO, and an unstable OO causes the odontoid-to-spinous process distance to decrease during extension (Lefebvre et al., 1993).
Fracture of the odontoid process: The odontoid process can fracture (Fig. 5-13), and any fracture of the odontoid is serious because of the close relationship between the odontoid process and spinal cord. Fracture of the odontoid superior to its attachment to the body of C2 is known as a type I fracture. Fracture along the attachment of the odontoid process to the body of C2 is known as a type II fracture, and fracture of the odontoid inferior to its attachment to the body of C2 that includes part of the body of C2 is known as a type III odontoid fracture. Type II fractures usually have the most difficulty reestablishing bony union at the fracture site, and surgical fusion generally is necessary in type II (Bednar, Parikh, & Hummel, 1995) and unstable type III fractures (Chiba et al., 1996). Type I fractures usually are treated with specialized plaster casts and cervical braces and stable type III fractures can be treated with halo braces (cervical stabilization braces that are attached by bone screws into the calvarium) (Chiba et al., 1996).
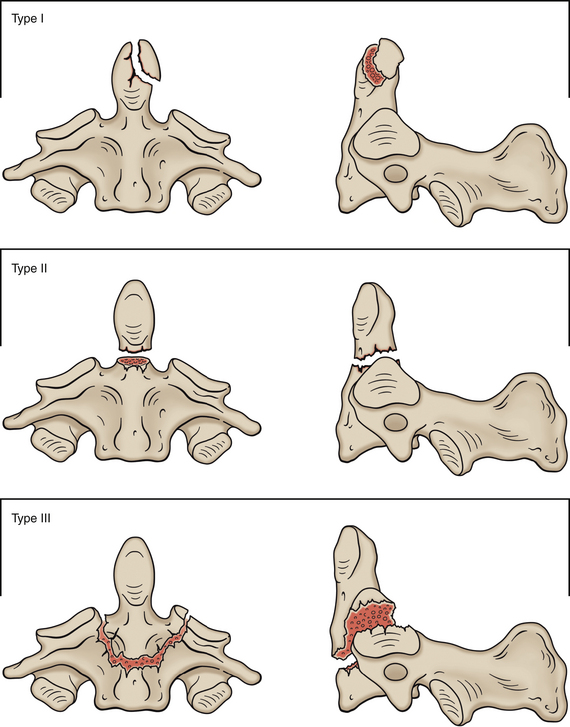
Body of the Axis
The body of C2 contains less cancellous bone than the dens (Standring et al., 2008). The body has a prominent inferior indentation and a subtle posterior one (Panjabi et al., 2001a). The anterior surface of the body is hollowed out because of the attachment of the longus colli muscle. As occurs throughout the cervical spine, the anterior longitudinal ligament attaches to the inferior border of the vertebral body of C2 in close association with the attachment of the anterior fibers of the anulus fibrosus. Another similarity of the inferior or discal border of C2 with the same border of the other cervical vertebrae is that its anterior aspect projects inferiorly. The posterior aspect of the vertebral body serves as an important attachment for the posterior longitudinal ligament and its superior continuation as the tectorial membrane. Specifically these structures attach to the posterior and inferior borders of the vertebral body. Also associated with the vertebral body is the first IVD of the spine, which is found between the inferior surface of the vertebral body of C2 and the superior surface of the vertebral body of C3.
Pedicles
The pedicles of the axis are thick from medial to lateral and from superior to inferior (see Fig. 5-12, B and E). Significant asymmetry between the left and right pedicles of C2 exists in the transverse (left to right) width and the angle formed between the C2 pedicles and vertebral body of C2 (Kazan et al., 2000). The left and right inferior vertebral notches of C2 are large, whereas the superior vertebral notches are almost nonexistent.
Superior Articular Processes
The superior articular processes of the axis can be thought of as smoothed out regions of the left and right pedicles of C2. That is, the superior articular processes do not project superiorly from the pediculolaminar junction, as occurs with the typical cervical vertebrae. Instead, they lie almost flush with the pedicle (see Fig. 5-12). This configuration, along with the very loose articular capsule at this level, allows much axial rotation (approximately 45 degrees unilaterally) to occur between C1 and C2. The articular cartilage of the superior articular process of C2 is convex superiorly, with a transverse ridge running from medial to lateral along the central region of the process. This ridge allows the anterior and posterior aspects of the facet to slope inferiorly, aiding in more effective rotation between C1 and C2 (Koebke & Brade, 1982) (see Atlanto-Axial Articulations). The articulation between the superior articular facet of C2 with the inferior articular facet of C1 is located anteriorly to the rest of the Z joints of the cervical spine. Therefore the superior articular processes of C2 and inferior articular processes of C1 are not a part of the articular pillars, as is the case with the lower cervical spine’s articular processes.
Laminae
The laminae of C2 are taller and thicker than those found in the rest of the cervical vertebrae. Because of the distinct architecture of the axis, the forces applied to it from above (by carrying the head) are transmitted from the superior articular processes to both the inferior articular processes and the vertebral body via the pedicle. Because the superior and inferior facets of the axis are arranged in different planes, the forces transmitted to the inferior articular processes are, by necessity, transferred through the laminae. This is accomplished by a rather complex arrangement of bony trabeculae (Pal et al., 1988). The laminae of the axis therefore are quite strong compared with the laminae of the rest of the cervical vertebrae.
Transverse Processes
The TPs of C2 are quite small and, like the TPs of C1 but unlike those of the rest of the cervical spine, do not possess distinct anterior and posterior tubercles. Developmentally they are considered to be homologues of the posterior roots or bars of the TPs, although minute homologues of the anterior tubercles are associated with the junction of the anterior aspect of the TPs with the vertebral body of C2.
The small left and right TPs of C2 face obliquely superiorly and laterally. Each has a foramen of the TP that, at C2, is an angular canal with two openings—one inferior and one lateral (Taitz, Nathan, & Arensburg, 1978). Therefore the vertebral artery courses laterally from the foramen of the TP of C2 to proceed to the more lateral foramen of the TP of C1.
Even though they are very small, the TPs of the axis serve as attachment sites for many muscles. Table 5-3 lists the muscles that attach to C2.
Table 5-3
Muscular Attachments to the Axis
| Region | Muscles Attached |
| Vertebral body | Longus colli |
| Transverse processes | Levator scapula Scalenus medius Splenius cervicis Intertransversarii (to upper and lower surfaces) |
| Spinous process | Obliquus capitis inferior Rectus capitis posterior major |
| Notch of spinous | Semispinalis cervicis Spinalis cervicis Interspinalis cervicis Multifidus (Also lamellar portion of ligamentum nuchae, when present) |
Spinous Process and Inferior Articular Processes
The spinous process of C2 is more prominently bifid than the other spinous processes of the cervical vertebrae because of the many muscles attaching to it (see Table 5-3). In 3% of the population the entire spinous process of C2 deviates slightly to the left or right side, and in approximately 39% of the population the superior-inferior lengths of the left and right sides of the bifid spinous processes are asymmetric. These asymmetries should be considered when viewing diagnostic images of, or when palpating, the C2 spinous process (Ji-Hong et al., 2010).
The inferior articular processes of C2 are typical for the cervical region. They arise from the junction of the pedicle and lamina and face anteriorly, inferiorly, and laterally.
Carotid Tubercles and Articular Processes of the Sixth Cervical Vertebra
Although the C6 vertebra is considered typical, its left and right anterior tubercles of the TPs are unique. These tubercles are prominent and are known as the carotid tubercles because each is so closely related to the overlying common carotid artery of the corresponding side. The common carotid artery may be compressed in the groove between the carotid tubercle and vertebral body of C6 (Standring et al., 2008).
In addition, the left and right articular pillars of C6 are unique to other cervical vertebrae in three ways. The first two features can be seen in the vertebrae of both children and adults, and the third feature is unique to adults only. First, each superior articular process and facet is distinctly more anterior in position than the inferior articular process and facet (rather than being almost “stacked” one above the other as found in C3-5). Second, the sulcus for the posterior primary division of C6 usually is a deep incisure that is distinct and can be seen from both lateral and posterior views of the vertebra. Finally, a prominent “muscular process” protrudes from the posterior aspect of adult C6 articular pillars. This process is found just inferior to the incisure for the posterior primary division (Herrera & Puchades-Ortis, 1998).
Vertebra Prominens (Seventh Cervical Vertebra)
The seventh cervical vertebra is known as the vertebra prominens because of its prominent spinous process (Fig. 5-14). The spinous process of C7 is the most prominent of the cervical region, although occasionally C6 is more prominent (C6 is the last cervical vertebra with palpable movement in flexion and extension). Also, the spinous process of T1 may be more prominent than that of C7 in some individuals.
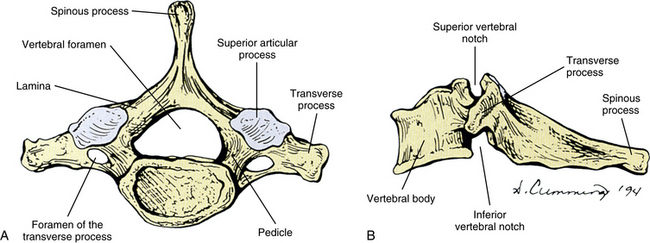
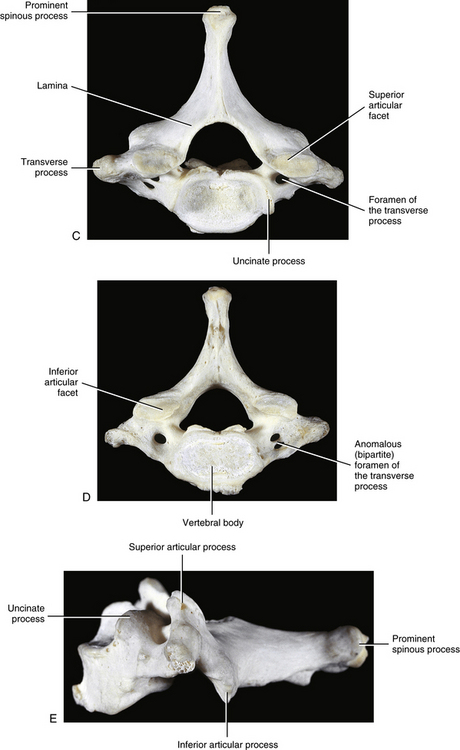
FIG. 5-14 A, Superior and, B, lateral views of the seventh cervical vertebra, the vertebra prominens. C, Superior, D, inferior, and, E, lateral views of the seventh cervical vertebra, the vertebra prominens.
The spinous process of C7 usually projects directly posteriorly. Unlike typical cervical vertebrae, the spinous process of C7 is not bifid. The funicular portion of the ligamentum nuchae attaches to the single posterior tip of the C7 spinous process. This ligament is discussed in more detail later in this chapter.
Because of its large spinous process and its location at the base of the neck, C7 serves as an attachment site for many muscles. Table 5-4 lists the muscular attachments of C7.
Table 5-4
| Region | Muscles Attached |
| Spinous process | Trapezius Rhomboid minor Serratus posterior superior Splenius capitis Spinalis cervicis Semispinalis thoracis Multifidus thoracis Interspinales |
| Transverse process | Middle scalene Spinalis capitis Intertransversarii Levator costarum (first pair) (Also suprapleural membrane [cupola]) |
| Articular process | Longissimus capitis |
| Vertebral body | Longus colli |
Transverse Processes
The TPs of C7 are also unique. The anterior tubercle of each C7 TP is small and short. The posterior tubercle is large, making the entire TP large. The anterior tubercle is the costal element of C7, and is unique because it develops from an independent primary center of ossification. This center usually unites with the TP by the fifth or sixth year of life. However, it may remain distinct and develop into a cervical rib. The formation of a cervical rib may also occur at C4 to C6 by the same mechanism, although this is less common. The intertubercular lamella usually is grooved by the ventral ramus of C7 anterior and lateral to the foramen of the TP (Standring et al., 2008). The suprapleural membrane, or cupola, which is the protective layer of connective tissue that reinforces the apical pleura of each lung, is attached to the posterior tubercle of the C7 TP.
Similar to the rest of the cervical region, the left and right C7 TPs contain a foramen. This foramen is usually the smallest of the cervical spine. Occasionally a double foramen is found in one of the TPs of C7 (Taitz, Nathan, & Arensburg, 1978) (Fig. 5-14, C and D). Frequently branches of the stellate ganglion run through the foramen of the TP of C7, although normally the only structures that course through this opening are accessory arteries and veins (Jovanovic, 1990). The accessory vessels comprise branches of the deep or ascending cervical arteries and their accompanying veins. The remainder of the C7 TP foramen is filled with areolar connective tissue. Recall that the vertebral artery and its associated sympathetic plexus run with the vertebral veins through the C6 foramen of the TP and the more superior vertebrae. Approximately 5% of the time, the vertebral artery and vein(s) traverse the foramen of the TP of C7 (Jovanovic, 1990).
Articulations of the Upper Cervical Region
The Z joints of the cervical region are covered earlier in this chapter. The atlanto-occipital and atlanto-axial joints are discussed here.
The articulations (joints) of the upper cervical spine are extremely important. These joints allow much of the flexion and extension that occurs in the cervical region and at least half of the axial (left and right) rotation of the cervical spine. In addition, the proprioceptive input from the atlanto-occipital and atlanto-axial joints, as well as proprioception from the suboccipital muscles, is responsible for the control of head posture (Panjabi et al., 1991).
Left and Right Atlanto-Occipital Articulations
The joints between the left and right superior articular surfaces of the atlas and the corresponding occipital condyles have been described as ellipsoidal (Standring et al., 2008) and condylar (Gates, 1980) in shape and type. The superior articular processes of the atlas are concave superiorly and face medially (see Fig. 5-10). Recall that the facets are narrow in their center, resulting in their peanut shape. The left and right facet surfaces are asymmetric 58% of the time (Pfirrmann et al., 2001). The occiput and the atlas are connected by articular capsules and the anterior and posterior atlanto-occipital membranes (see Figs. 5-16 and 5-21). The fibrous capsules surround the occipital condyles and the superior articular facets of the atlas. Synovial folds usually (98% of joints in children and 55% in adults) extend from the joint capsules into these joints (Tang et al., 2007). These synovial folds are largest in the anterolateral aspects of the joints (Tang et al., 2007), even though the joint capsules are thickest posterolaterally. Smaller intraarticular “fat pads” have also been identified in these joints. The fat pads generally extend from the articular capsule into the medial and lateral aspects of narrow joint “waist,” approximately midway along the anterior-posterior distance of the joint (Mercer & Bogduk, 1993). Each capsule is further reinforced in the posterolateral region by a ligamentous band that passes between the jugular processes of the occiput and the lateral mass of the atlas. This band has been called the lateral atlanto-occipital ligament (Oliver & Middleditch, 1991). The atlanto-occipital joint capsules are thin and sometimes completely nonexistent medially. When present, this medial deficiency frequently allows the synovial cavity of the atlanto-occipital joint to connect with the bursa or joint cavity between the dens and the transverse atlantal ligament (Cave, 1934; Standring et al., 2008).
More than 2 mm of anterior-to-posterior translation of the occiput on the atlas (as measured from the anterior rim of the foramen magnum, or basion, to the posterior aspect of the anterior arch) on flexion and extension lateral radiographs is considered to be related to instability of this region (Hankinson & Anderson, 2010). However, the value for the lower limit of normal can be considered slightly higher in children and translation of less than 3 mm should not be considered as serious in children with Down syndrome (Matsuda et al., 1995). In addition to ligamentous laxity, atlanto-occipital instability can be due to a superior articular facet of the atlas that has a relatively hoizontal orientation (as compared with the normal concave configuration). The normally convex occipital condyles may also be more horizontally oriented (i.e., “flattened”) in atlanto-occipital instability (Hankinson & Anderson, 2010).
Atlanto-Axial Articulations
The atlas and axis articulate with one another at three synovial joints: two lateral joints and a single median joint complex (Fig. 5-15, B).
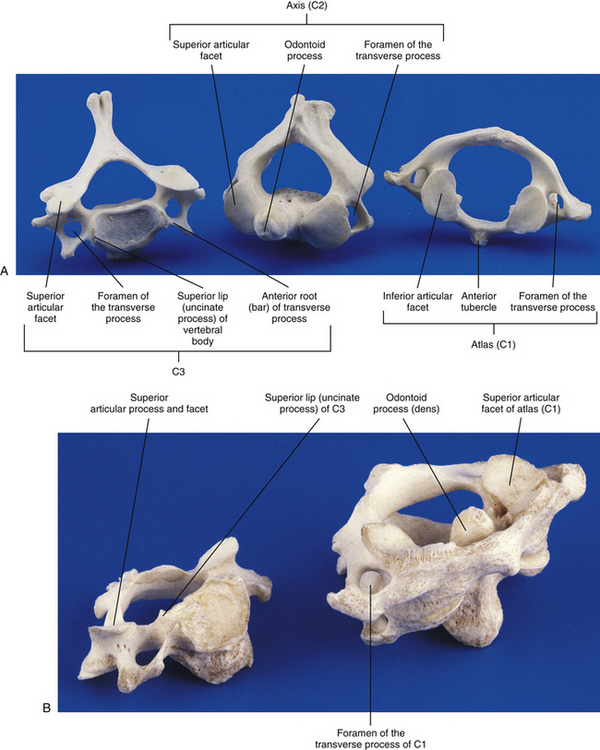
FIG. 5-15 A, Inferior view of the atlas and a superior view of the axis and third cervical vertebra. B, Atlas and axis in typical anatomic relationship, with C3 to the side.
Lateral Atlanto-Axial Joints
The lateral atlanto-axial joints are planar joints that are oval in shape. The bony surfaces of inferior articular process of the atlas and the superior articular process of the axis are generally flat (Cattrysse et al., 2008); however, the articular cartilage covering these bony surfaces results in a slight convexity of both articular surfaces in the living. The inferior articular facet of the atlas is usually slightly larger than the superior articular facet of the axis. Asymmetry between the shapes of the left and right articular facets constituting these joints and asymmetry of the plane of articulation of the left and right lateral atlanto-axial joints are common (Ross, Bereznick, & McGill, 1999; Meseke, Duray, & Brillon, 2008). The fibrous capsule of each lateral joint is thin and loose and attaches to the outermost rim of the articular margins of the atlas and axis (inferior articular facet of the atlas and superior articular facet of the axis). Each capsule is lined by a synovial membrane. Synovial folds, originating from the articular capsule, usually (80% of joints in children and 85% in adults) protrude into the lateral atlanto-axial joints. These folds are most prominent in the anterolateral aspects of the joints (Mercer & Bogduk, 1993; Tang et al., 2007) (see previous section, Articular Processes and Zygapophysial Joints for further detail on synovial folds in these joints). A posteromedial accessory ligament attaches inferiorly to the body of the axis near the base of the dens and courses superiorly to the lateral mass of the atlas near the attachment site of the transverse ligament. This ligament is known as the accessory atlanto-axial ligament (see Ligaments of the Cervical Region).
Median Atlanto-Axial Joint
The median atlanto-axial joint is a pivot (or trochoid) joint between the dens and a ring of structures that encircles the dens. These structures are the anterior arch of the atlas anteriorly and the transverse ligament posteriorly (see Cruciform Ligament and Fig. 5-19 later in this chapter). The hyaline articular cartilages on the atlas and the odontoid process, comprising the median atlanto-axial joint, are oval, but the facet on the odontoid process has its longest axis oriented longitudinally and that of C1 oriented horizontally. The posterior facet of the dens (that articulates with the transverse ligament of the atlas) is also oval and is oriented longitudinally. The area of the articular cartilage of the atlas is 58.2 mm2, and that of the anterior surface of the dens is 55.1 mm2. The thickness of the articular cartilage on C1 and both the anterior and posterior articular surfaces of the odontoid process is approximately 0.80 mm (Ebraheim et al., 1997a).
The anterior and posterior components of the median atlanto-axial articulation both have synovial-lined fibrous capsules forming distinct joint cavities. The synovial-lined capsules attach to the outer edge of the articular cartilages of the opposing surfaces, and the capsules have slack in them to allow for significant movement (Ebraheim et al., 1997a). The anterior portion of the joint usually (80% of children and 70% of adults) contains a synovial fold that descends into the superior half of the joint from the superior aspect of the joint capsule. This fold may help to lubricate the joint (as is the case with all Z joint synovial folds) and may also prevent bone-on-bone compression during the large amount of axial rotation that occurs at this articulation (Tang et al., 2007). The posterior joint cavity is the larger of the two, and sometimes it has a bursa associated with it. In either case it is located between the anterior surface of the transverse ligament and the posterior grooved surface of the odontoid process. This posterior joint is often continuous with one of the atlanto-occipital joints (Standring et al., 2008).
Clinical Applications Related to the Atlas and Axis
Of patients with rheumatoid arthritis, 86% have involvement of the cervical region and the median atlanto-axial joint is affected in approximately 20% to 25% of these patients, although the incidence may be higher (Lehto et al., 2010). Somewhat similar involvement is seen in patients with psoriatic arthritis (35% to 75% cervical involvement, 10% upper cervical involvement) (Ouedraogo et al., 2007). Pathologic horizontal displacement of the atlas on the axis occurs in a significant number of the patients with upper cervical involvement. Such displacements are caused by erosive synovitis of the capsules, extending into the joints and affecting the articular cartilages, including the fibrocartilage of the transverse ligament of the atlas. (The erosions of the articular cartilages and other pathologic changes can be seen on CT and MRI.) If the cartilage and adjacent bone become significantly eroded, the anterior arch of the atlas can displace slightly inferiorly on the odontoid process, a condition known as atlanto-axial impaction. Atlanto-axial impaction may help to stabilize anterior atlanto-axial subluxation, but it can be painful when the individual is in the upright position (the pain usually diminishes during bed rest) (Lehto et al., 2010). In addition, approximately 7% of patients with rheumatoid arthritis have a small vertical subluxation (i.e., ascension of the odontoid process toward the foramen magnum), although symptoms related to these relatively subtle vertical displacements in patients with rheumatoid arthritis are rare (Ebraheim et al., 1997a).
Rheumatoid arthritis can also affect the atlanto-occipital and lateral atlanto-axial joints, and when severe can lead to destructive changes and/or ankylosis (fusion) of these joints (Iizuka et al., 2009; Nagayoshi et al., 2009). Signs and symptoms of myelopathy secondary to horizontal subluxations of the atlas on the axis (attributable to pathology of the median atlanto-axial joint) or ankylosis of the upper and lower cervical synovial joints are relatively frequent and ankylosis of the atlanto-occipital or lateral atlanto-axial joints has been associated with severe cervical myelopathy below the level of C2 (Iizuka et al., 2009).
Also, degenerative changes of the median atlanto-axial joint in individuals without rheumatoid arthritis can begin at 50 years of age and can progress to completely obliterate the anterior and posterior joint spaces (Ebraheim et al., 1997a; Milz et al., 2001). The degenerative changes can also result in osteophytes that develop on the superior aspect of either the atlas or the odontoid process (or both). In some instances large osteophytes from the atlas may abut the occiput and be related to a third occipital condyle and may or may not create a median atlanto-occipital joint (von Ludinghausen et al., 2005).
Chronic atlanto-axial instability has been related to the development of a fibrous mass, called a retro-odontoid pseudotumor, immediately posterior to the odontoid process. The incidence of retro-odontoid pseudotumor is higher in patients with ossification of the anterior longitudinal ligament (Chikuda et al., 2009).
Children frequently have increased ligamentous laxity, which leads to increased spinal motion. This occurs most often in the cervical region. Increased motion seen during physical or x-ray examination should be differentiated from pathologic subluxation (Fesmire & Luten, 1989). For example, the space between the anterior arch of the atlas and odontoid process, known as the predental space or atlanto-odontal interspace, usually does not exceed 3 mm. A predental space greater than 3 mm generally is considered to indicate a tear of the transverse ligament (see later discussion) or pathologic subluxation of C1 on C2. However, a 3-mm or greater predental space has been found in 20% of healthy patients less than 8 years of age (Fesmire & Luten, 1989). Although a space of greater than 3.5 mm usually is considered abnormal in children, spaces of up to 5 mm have been seen in normal children (Fesmire & Luten, 1989). Therefore x-ray evaluation of atlanto-axial stability in children should be tempered with sound clinical judgment.
Pathologic subluxation of the atlas on the axis resulting in compromise of the spinal cord (compressive myelopathy) has been associated not only with rheumatoid arthritis (Kaufman & Glenn, 1983) but also less frequently with ankylosing spondylitis. Displacement of the atlas on the axis also has been found in 9% of 5- to 21-year-old patients with Down syndrome. In addition, significant degeneration of the entire cervical spine with osteophyte formation, narrowing of foramina, and narrowing of the disc space has been found with increasing incidence in the adult Down syndrome population. The premature aging process that occurs in individuals with this syndrome may be one possible explanation for this latter finding (Van Dyke & Gahagan, 1988).
Ligaments of the Cervical Region
The ligaments of the cervical region can be divided into upper and lower cervical ligaments. The upper ligaments are those associated with the occiput, atlas, and the anterior and lateral aspect of the axis. The lower cervical ligaments encompass all other ligaments of the cervical region. The ligaments of both categories are discussed in the following sections. The points of insertion and the function of each are discussed, beginning with those located most posteriorly and progressing to those located most anteriorly. Tables 2-2 and 2-3 summarize the spinal ligaments.
Upper Cervical Ligaments
Table 2-2 summarizes the upper cervical ligaments.
Posterior Atlanto-Occipital Membrane
The posterior atlanto-occipital membrane is a rather thin structure that attaches to the posterior arch of the atlas and the posterior rim of the foramen magnum (Fig. 5-16). It is usually closely adherent to the posterior aspect of the spinal dura mater, and together these structures have been called the posterior atlanto-occipital membrane–spinal dura complex (Hack et al., 1995a). The posterior atlanto-occipital membrane functions to limit flexion of the occiput on the atlas. The ligament is so broad from left to right that the term membrane applies. It spans the distance between the left and right lateral masses. Laterally this ligament arches over the left and right grooves for the vertebral artery on the posterior arch of the atlas (see Fig. 5-16). This allows passage of the vertebral artery, vertebral veins, and the suboccipital nerve. These structures are covered in more detail later in this chapter. This lateral arch of the posterior atlanto-occipital membrane occasionally ossifies, creating a posterior ponticle. A posterior ponticle, in turn, creates a foramen for the previously mentioned vascular and neural structures. (See previous section on the atlas for elaboration on posterior ponticle and the arcuate, or arcual, foramen both created by the ossified posterior atlanto-occipital membrane.)
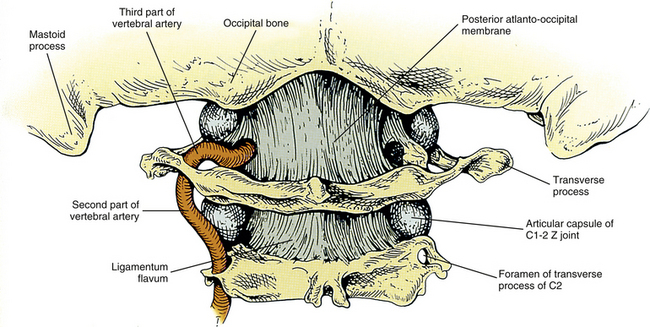
Tectorial Membrane
The tectorial membrane is the superior extension of the posterior longitudinal ligament (Fig. 5-17). It begins by attaching to the posterior aspect of the vertebral body of C2. It then crosses over the odontoid process and inserts onto the anterior rim of the foramen magnum (specifically the upper region of the basilar part of the occipital bone, or clivus). The tectorial membrane has superficial and deep fibers. The deep fibers have a median band that extends all the way to the basilar portion of the occipital bone. Two lateral bands of deep fibers pass medially to the atlanto-occipital joints before attaching to the occiput. The superficial fibers extend even more superiorly than the deep fibers and blend with the cranial dura mater at the upper region of the basilar part of the occipital bone (clivus). This ligament limits both flexion and extension of the atlas and occiput (Standring et al., 2008).
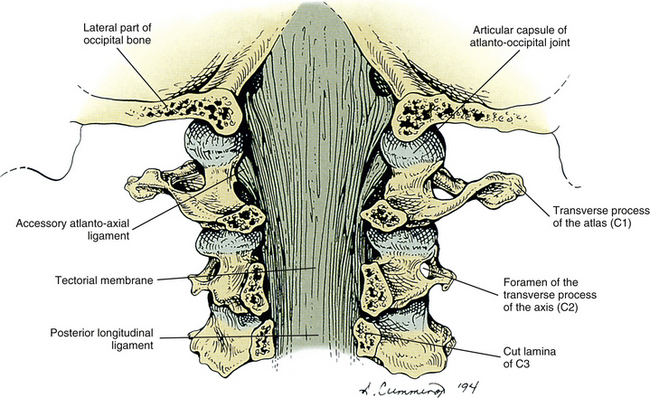
Accessory Atlanto-Axial Ligaments
Each of the accessory atlanto-axial ligaments (left and right) courses from the base of the odontoid process to the inferomedial surface of the lateral mass of the atlas on the same side (Figs. 5-17 and 5-18). They help to strengthen the posteromedial aspect of the capsule of the lateral atlanto-axial joints. They are considered to be deep fibers of the tectorial membrane.

FIG. 5-18 Anterior aspect of the vertebral canal and foramen magnum as seen posteriorly. The tectorial membrane has been removed, and many of the upper cervical ligaments can be seen. Notice the centrally located cruciform ligament with its narrow superior and inferior longitudinal bands and its stout transverse ligament. The alar and accessory atlanto-axial ligaments also can be seen.
Cruciform Ligament
The cruciform ligament is named on the basis of its cross shape. Actually, it may be divided into several parts: a large transverse ligament, a superior longitudinal band, and an inferior longitudinal band (Fig. 5-18). Each portion is discussed next.
Transverse ligament: The transverse ligament of the atlas (TLA), or transverse portion of the cruciform ligament, has been called the most important ligament of the occiput–C1-C2 complex of joints (White & Panjabi, 1990). It is a strong ligament that runs from a small medial tubercle of one lateral mass of the atlas (colliculus atlantis, Fig. 5-10, G) to the same tubercle on the opposite side. The TLA lies in the horizontal plane. However, approximately a 21-degree angle with the frontal (coronal) plane is created from the origin of the transverse ligament to the region where it passes behind the odontoid process (Panjabi, Oxland, & Parks, 1991a). It courses approximately 5 mm inferior to the odontal attachments of the apical odontoid ligament and the left and right alar ligaments (Ebraheim et al., 1997a). The superior-inferior width of this ligament is greatest at its center, where it passes posterior to the odontoid process. Anteriorly the TLA is lined by a thin layer of fibrocartilage (Ebraheim et al., 1997a), which enables the TLA to form a diarthrodial joint with the odontoid as it passes posterior to this structure.
The TLA allows the atlas to pivot on the axis (Fig. 5-19). It also holds the atlas in its proper position, thereby preventing compression of the spinal cord during flexion of the head and neck. Because the transverse ligament fits into the groove on the posterior surface of the odontoid, it holds the atlas in proper position even when all other ligaments are severed (Standring et al., 2008). The atlanto-odontal interspace can increase to as great as 8 mm (normally 3 mm) when the TLA is severed (Menezes & Traynelis, 2008). Panjabi and colleagues (1991a) found that this ligament appears to exist in two distinct layers, superficial and deep.
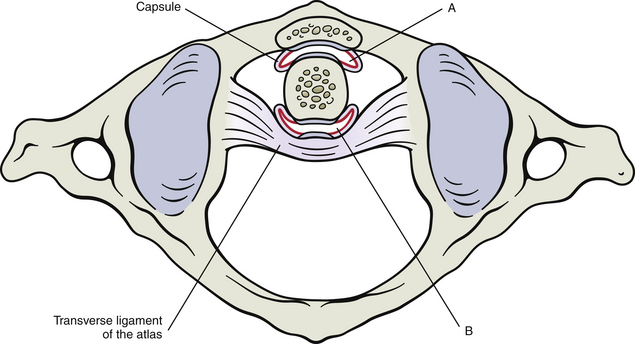
FIG. 5-19 Median atlanto-axial joint. Notice the role played by the transverse ligament of the atlas in allowing the atlas to pivot around the odontoid process. The two parts of the joint are distinct, each with its own joint capsule (Ebraheim et al., 1997a). The synovial lining of the capsules is shown in red. A, Anterior part of the joint, between the anterior arch of the atlas and the odontoid. B, Posterior part of the joint, between the odontoid process and the transverse ligament of the atlas.
Fibrocartilage frequently develops in regions of ligaments that are constantly compressed against bone. Examples of this include portions of the Achilles tendon, tendons of the rotator cuff, and extensor tendons of the fingers and toes. Similarly, the region of the TLA that passes posterior to the odontoid process also develops fibrocartilage. However, this region is unique in that a fibrous capsule surrounds the fibrocartilage facet on the transverse ligament, forming a diarthrodial (synovial) joint with the posterior articular facet of the odontoid process. The fibrocartilage of the TLA possesses substances also found in typical articular cartilage (e.g., link protein, aggrecan, and type II collagen). Some cases of rheumatoid arthritis are related to an autoimmune response to these substances. Such autoimmune responses could erode the TLA, resulting in the upper cervical instability seen in a significant number of patients with rheumatoid arthritis. Furthermore, fibrocartilage has less ability to repair than typical ligaments, complicating the healing of erosions affecting the TLA. This slower rate of healing may help to explain why calcium deposition or ossification frequently is seen in TLAs of arthritic median atlanto-axial joints (Milz et al., 2001).
Superior longitudinal band: The superior longitudinal band of the cruciform ligament runs from the transverse ligament to the anterior lip of the foramen magnum (see Fig. 5-18). More specifically, it attaches to the superior aspect of the basilar part of the occipital bone (clivus). It is interposed between the apical ligament of the odontoid process, which is anterior to it, and the tectorial membrane, which is posterior to it. Although the superior longitudinal band may limit both flexion and extension of the occiput, its primary function may be to hold the transverse ligament in its proper position, thus aiding the transverse ligament in holding the atlas against the odontoid process.
Inferior longitudinal band: The inferior longitudinal band of the cruciform ligament attaches the TLA to the body of C2, preventing the transverse ligament from riding too far superiorly (see Fig. 5-18). It also helps to limit flexion of the occiput and atlas on the axis (with the aid of the superior band and transverse ligament).
Alar Ligaments
The left and right alar ligaments originate from the posterior and lateral aspect of the odontoid process with some of the fibers covering the entire posterior surface of the dens (Panjabi et al., 1991a). Each alar ligament passes laterally to insert onto a roughened region of the medial surface of the occipital condyle of the same side (Fig. 5-20; see also Fig. 5-18), although the superior to inferior direction of the ligaments varies with some ligaments having an inferior to superior course (caudocranial orientation, 44%), some having a horizontal course (49.5%), and rarely a craniocaudal course may exist (6.5%). This variation from caudocranial to craniocaudal course is generally dependent upon the length of the odontoid process, with the course becoming more horizontal and then craniocaudal with odontoid processes of increasing length. The average angle between the left and right alar ligaments is 140 to 180 degrees. The left and right alar ligaments are usually (88%) asymmetric in their relationship to the coronal and horizontal planes (Pfirrmann et al., 2001). The alar ligaments are approximately the width of a pencil and are strong.
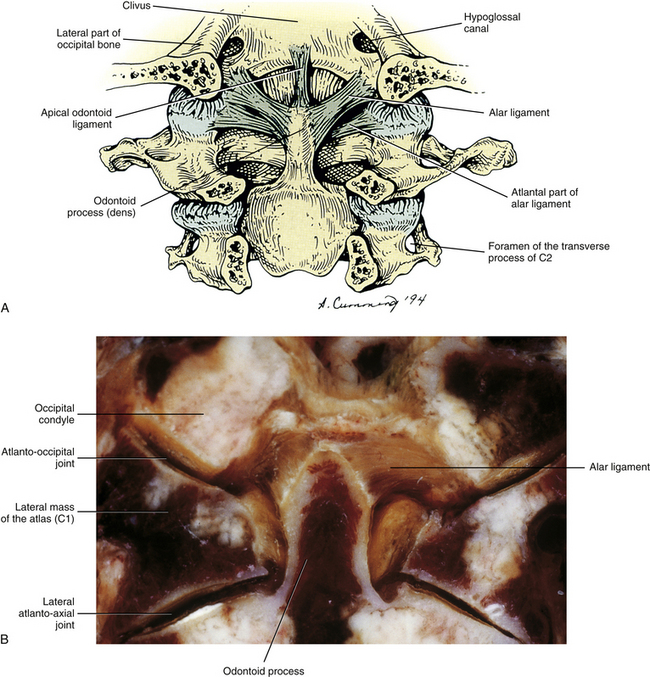
FIG. 5-20 A, Alar and apical odontoid ligaments. This is the same view as that of Figures 5-17 and 5-18. The tectorial membrane and cruciform ligament have been removed. Notice that some fibers of each alar ligament attach to the lateral mass of the atlas. These fibers have been described by Dvorak and Panjabi (1987). B, Coronal section of upper cervical region demonstrating the alar ligaments.
The functions of the alar ligaments are complex and are not understood completely. However, each alar ligament limits contralateral axial rotation (Dvorak & Panjabi, 1987). For example, the left alar ligament primarily limits right rotation. More specifically, the fibers of the left alar ligament, which attach to the odontoid process posterior to the axis of movement, act in concert with those fibers of the right alar ligament, which attach to the odontoid in front of the axis of movement. Both of these segments of the alar ligaments act together to limit right axial rotation. The opposite is also true: Right posterior odontal fibers and left anterior odontal fibers limit left rotation (Standring et al., 2008). Because the alar ligaments limit or check rotation, they are also known as the check ligaments.
The alar ligaments also limit flexion of the upper cervical spine after the tectorial membrane and cruciform ligaments have torn. The alar ligaments themselves are most vulnerable to tearing during the combined movements of axial rotation and flexion. This combination of movements may occur during a motor vehicle accident (e.g., frontal collision while looking in the rearview mirror) (Foreman & Croft, 1992). Injury as a result of this same pair of movements also can irreparably stretch the alar ligaments while sparing the cruciform ligament. When an alar ligament is torn or stretched, increased rotation occurs at the atlanto-occipital and atlanto-axial joint complexes, and increased lateral displacement occurs between the atlas and the axis during lateral flexion (Dvorak & Panjabi, 1987).
Dvorak and Panjabi (1987) found that in addition to attaching to the occipital condyle of the same side, a portion of each alar ligament usually attaches to the lateral mass of the atlas on the same side as well (see Fig. 5-19, A). They also occasionally found fibers coursing from the odontoid process to the anterior arch of the atlas. They named these latter fibers the anterior atlanto-dental ligament and believed that this ligament, when present, gives functional support to the transverse ligament. They stated that the alar ligaments also help to limit lateral flexion at the atlanto-occipital joint. The atlantal fibers of the alar ligament on the side of lateral flexion tighten first during this motion, followed by tightening of the occipital fibers of the alar ligament on the opposite side.
A ligamentous band that attaches near the left and right occipital condyles immediately posterior to the attachment of the alar ligaments has been identified in 10% - 77.8% of cadavers (Tubbs et al., 2010). The band has been called the transverse occipital ligament and usually attaches to the tip of the odontoid process between the superior longitudinal band of the cruciform ligament and the apical odontoid ligament and often has connections with the alar ligaments. The band may function to help reinforce the alar ligaments or the superior longitudinal band of the cruciform ligament (Tubbs et al., 2010).
Apical Ligament of the Odontoid Process
The apical ligament of the odontoid process is thin, is approximately 2.5 cm in length, and runs from the posterior and superior aspects of the odontoid process to the anterior wall of the foramen magnum (inferior aspect of the clivus) (see Fig. 5-19, A).
The supraodontoid region in which this ligament resides has been referred to as the “apical cave.” This region contains a coiled venous plexus, two adipose tissue pads (each approximately 1 cm in diameter), branches of the C1 and C2 recurrent meningeal nerves, and the apical odontoid arterial arcade (Haffajee, Thompson, & Govender, 2008).
The insertional fibers of the apical odontoid ligament blend with the deep fibers of the superior longitudinal band of the cruciform ligament. Its course from the odontoid to the clivus results in approximately a 20-degree anterior tilt of the apical odontoid ligament. Its insertion is wider than its origin, giving it a V shape (Panjabi et al., 1991a). Embryologically this ligament develops from the core of the centrum of the proatlas (see Chapter 12) and contains traces of the notochord (Standring et al., 2008). The apical odontoid ligament probably functions to prevent some vertical translation and anterior shear of the occiput (Panjabi et al., 1991a).
Anterior Atlanto-Occipital Membrane
The anterior atlanto-occipital membrane is located in front of the apical odontoid ligament and courses from the superior aspect of the anterior arch of the atlas to the anterior margin of the foramen magnum (Fig. 5-21). It is composed of densely woven fibers and is so broad that it can best be described as a membrane. The anterior atlanto-occipital membrane blends laterally with the capsular ligaments of the atlanto-occipital articulation (see Fig. 5-21). It functions to limit extension of the occiput on C1. Fibers continuous with the anterior longitudinal ligament strengthen the anterior atlanto-occipital membrane medially and form a tough central band between the anterior tubercle of the atlas and the occiput (Standring et al., 2008) (see Fig. 5-21). The anterior longitudinal ligament is discussed in more detail in the following section.
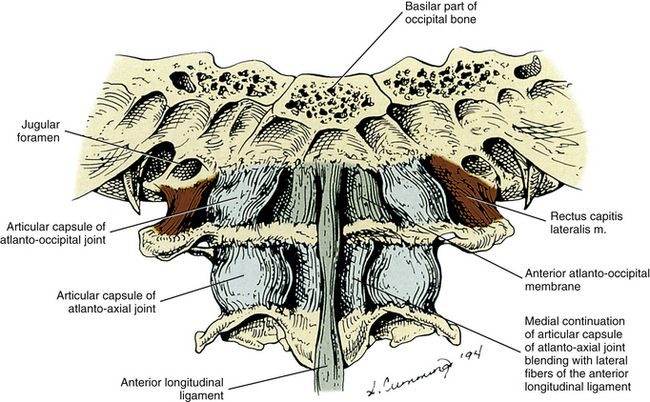
FIG. 5-21 Anterior view of the occiput, atlas, axis, and related ligaments. The anterior longitudinal ligament narrows considerably between the atlas and occipital bone and blends laterally with the anterior atlanto-occipital membrane. The articular capsules of the atlanto-occipital and lateral atlanto-axial joints also are seen clearly in this figure.
Lower Cervical Ligaments
Table 2-3 summarizes the lower cervical ligaments.
Anterior Longitudinal Ligament
The anterior longitudinal ligament (ALL) is wide and covers the anterior aspect of the vertebral bodies and IVDs from the occiput to the sacrum. Superiorly the ALL thickens medially to form a cord that attaches to the body of the axis and the anterior tubercle of the atlas (see Fig. 5-21). Some of the atlantal fibers diverge laterally as the ALL fibers attach to the inferior aspect of the anterior arch of the atlas. Further superiorly the ALL becomes continuous with the medial portion of the anterior atlanto-occipital membrane (see previous discussion). The ALL is approximately 3.8 mm wide at C1-2, is somewhat wider at C2-3, and increases in width to 7.5 mm from C3 to T1. The ALL is firmly attached to a significant portion of the superior and inferior bony end plates in all regions of the spine. However, regional differences exist for the ALL attachment sites. The ALL is firmly attached to the central region of the vertebral bodies 40% of the time in the cervical region, but is firmly attached to the center of the thoracic or lumbar vertebral bodies only rarely. In contrast, the ALL is almost never firmly attached to the IVD in the lumbar region and is firmly attached to this structure infrequently in the cervical region (20% of the time); however, the ALL is firmly attached to the IVD about half the time (50%) in the thoracic region (Cramer et al., 1996, 1998). Laterally the ALL is sometimes difficult to distinguish from the anterolateral fibers of the anulus fibrosus.
Several layers are associated with the ALL. The superficial fibers span several vertebrae, whereas the deep fibers course from one vertebra to the next. This ligament tends to be thicker from anterior to posterior in the regions of the vertebral bodies rather than the areas over the IVDs. Therefore the ALL helps to smooth the contour of the anterior surface of the vertebral bodies by filling the natural concavity of the anterior vertebral bodies. The ALL functions to limit extension and frequently is damaged in extension injuries to the cervical region (Halliday et al., 1964; Bogduk, 1986a). However, during extension injuries to the spine, fibers of the anterior anulus fibrosus of the IVDs frequently tear before those of the more anteriorly located ALL because the fibers of the anulus fibrosus are shorter than those of the ALL and reach their failure loads before the fibers of the ALL (Taylor, 1999).
The ALL can ossify (OALL). OALL reduces cervical mobility and has been associated with the presence of a fibrous mass directly posterior to the odontoid process, known as retro-odontoid pseudotumor (see preceding section titled Median Atlanto-Axial Joint) (Chikuda et al., 2009).
Posterior Longitudinal Ligament
The posterior longitudinal ligament (PLL) is the inferior continuation of the tectorial membrane (see Figs. 5-17 and 5-22). It courses from the posterior aspect of the body of C2, inferiorly to the sacrum, and possibly to the coccyx (Behrsin & Briggs, 1988). The ALL and PLL have similar tensile properties (Przybylski et al., 1996). That is, they can withstand similar loads applied to the spine, although the ALL limits forces applied in extension and the PLL resists forces applied in flexion. The PLL is wide and regularly shaped in the cervical and upper thoracic regions and is also three to four times thicker, from anterior to posterior, in the cervical region than in the thoracic or lumbar regions (Bland, 1989). Its superficial fibers span several vertebrae, and its deep fibers course between adjacent vertebrae. Panjabi and colleagues (1991b) found the cervical PLL to be firmly attached to both the vertebral bodies and the IVDs, whereas Bland (1989) found the PLL to have a stronger discal attachment. In either case, the PLL probably functions to help prevent posterior IVD protrusion. Although the PLL is attached to the entire length of the vertebral bodies in the cervical region (Przybylski et al., 1998), it is more loosely attached to the central region of the vertebral bodies to allow the exit of the basivertebral veins from the vertebral bodies (Standring et al., 2008). The PLL in the middle and lower thoracic and lumbar regions differs from the PLL in the cervical region in that it becomes narrow over the vertebral bodies and then widens considerably over the IVDs in the thoracic and lumbar areas.
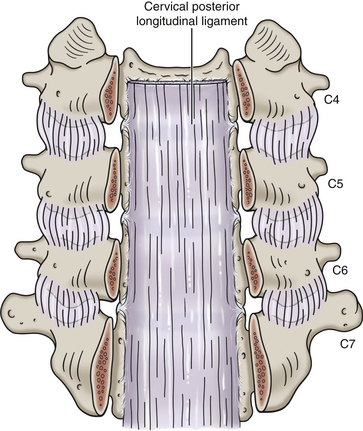
FIG. 5-22 Cervical posterior longitudinal ligament (PLL). The PLL in the cervical region is different than the PLL in the thoracic and lumbar regions. The PLL in the cervical region is uniformally wide throughout the cervical region, not only behind (posterior to) the intervertebral discs but also the vertebral bodies. The PLL in the thoracic and lumbar regions is narrow as it passes behind each vertebral body and then fans out to become very wide as it attaches to each intervertebral disc.
The PLL receives a significant nociceptive and vasomotor innervation. The nociceptive innervation may make this ligament one of the most pain sensitive of the spine. The vasomotor fibers probably help to increase regional blood flow to promote healing after ligamentous damage (Imai et al., 1997).
The PLL occasionally ossifies. This occurs most frequently in the cervical region (Fig. 5-23) and occasionally occurs in the lumbar region. (Do not confuse this with ossification of the ligamenta flava, which occurs most frequently in the thoracic region.) Ossification of the PLL (OPLL) is clinically relevant because it may be a source of compression of the spinal cord (even with relatively minor trauma) in the cervical region, and has been associated with radicular symptoms in the lumbar region (Hasue et al., 1983; Chikuda et al., 2011). OPLL is found primarily in the middle or lower cervical spine of middle-aged and elderly men (almost 3.5:1 more than women) of Japanese origin, living in Japan (Yamada et al., 2003). Japanese people also have a higher incidence of ossification of other spinal ligaments than other ethnic populations. The incidences of OPLL, ossification of the ALL, and ossification of the ligamentum nuchae in Japanese men 48 to 57 years old are 4.1%, 23.1%, and 23.3%, respectively (Shingyouchi, Nagahama, & Niida, 1996). A close association of OPLL and diffuse idiopathic skeletal hypertrophy (DISH) has also been found, and OPLL is now considered to be a variant of DISH (Yamada et al., 2003). DISH has been associated with obesity and glucose intolerance (Shingyouchi, Nagahama, & Niida, 1996). Although the etiology of OPLL is unknown and undoubtedly complex, mechanical stress, genetic factors, dietary factors, and vitamin K2 (menaquinone) metabolism have all been associated with this condition (Matsunaga et al., 1996; Shingyouchi, Nagahama, & Niida, 1996; Yamada et al., 2003). Estrogen levels also play a role in OPLL. (Estrogen promotes osteoblastic activity.) With respect to diet, OPLL and ossification of the ALL have been significantly linked to obesity and diabetes or impaired glucose tolerance, whereas ossification of the ligamentum nuchae (OLN) has been linked to obesity only (Shingyouchi, Nagahama, & Niida, 1996). Furthermore, OPLL also may be associated with the high-salt, low-meat diets of Japanese and Taiwanese (Wang et al., 1999). Hypertrophy of the PLL (HPLL) is a distinct condition that is a pathologic thickening of the PLL, and HPLL may or may not be a precursor to OPLL. Both HPLL and OPLL show unique cellular characteristics. Cells of both HPLL and OPLL ligaments seem to possess similar regulatory mechanisms (perhaps secreting a similar growth factor) that may allow for accelerated growth and division of cells in the PLL (Motegi et al., 1998).
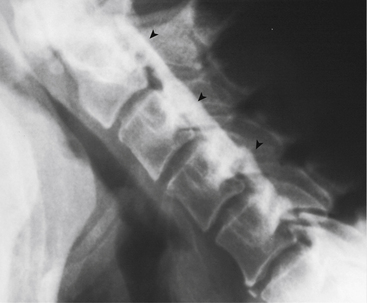
FIG. 5-23 Lateral cervical flexion x-ray showing ossification (arrowheads) of the posterior longitudinal ligament (OPLL). The OPLL is related to diffuse idiopathic skeletal hyperostosis (DISH). The ossified PLL is dramatically narrowing the vertebral canal (cervical vertebral canal [spinal] stenosis), which can cause myelopathy of the cervical spinal cord. See text for further details. (Courtesy Dr. William Bogar, National University of Health Sciences, Lombard, IL.)
Ligamenta Flava
The ligamenta flava (singular, ligamentum flavum) are paired ligaments (left and right) that run between the laminae of adjacent vertebrae (see Fig. 5-16). They are found throughout the spine beginning with C1-2 superiorly and ending with L5-S1 inferiorly. The posterior atlanto-occipital membrane is the homologue of the ligamenta flava at the level of occiput-C1. Each ligamentum flavum is approximately 5 mm thick from anterior to posterior (Panjabi et al., 1991b). These ligaments are thinnest in the cervical region, become thicker in the thoracic region, and are thickest in the lumbar region.
Each ligament passes from the anterior and inferior aspect of the lamina of the vertebra above to the posterior and superior aspect of the lamina of the vertebra below. The ligamenta flava increase in length from C2-3 to C7-T1. This implies that the distance between the laminae also increases in a similar manner. Laterally each ligamentum flavum helps to support the anterior aspect of the Z joint capsule. Although each ligament is considered to be distinct, a ligamentum flavum frequently blends with the ligamentum flavum of the opposite side (Panjabi et al., 1991b) and also blends with the interspinous ligament. Small gaps exist between the left and right ligamenta flava, allowing for the passage of veins that unite the posterior internal (epidural) vertebral venous plexus with the posterior external vertebral venous plexus. The ligamentum flavum between C1 and C2 is usually thin and membranous and is pierced by the C2 spinal nerve. In fact, Panjabi and colleagues (1991b) were unable to find ligamenta flava between C1 and C2 in their study of six cervical spines.
The ligamentum flavum is unique in that it contains yellow elastin, which causes it to constrict naturally. Therefore this ligament actually may do work; that is, it may aid in extension of the spine. It also slows the last few degrees of spinal flexion. However, the most important function of the elastin may be to prevent buckling of the ligamentum flavum into the spinal canal during extension.
The ligamentum flavum may undergo degeneration with age or after trauma. Under such circumstances, it usually increases in thickness and may calcify or become infiltrated with fat (Ho et al., 1988). These changes may cause the ligament to lose its elastic characteristics, which can result in buckling of the thickened ligamentum flavum into the vertebral canal or medial aspect of the IVF. The buckling further results in narrowing of these regions, which can compromise the neural elements running within them (e.g., spinal cord, cauda equina [lumbar region], or exiting nerve roots). Ossification of the ligamentum flavum is reported to occur most often in the thoracic and thoracolumbar regions of the spine, where it may compress either the posterior aspect of the spinal cord or the exiting nerve roots (Hasue et al., 1983) (see Chapter 6).
Interspinous Ligaments
The interspinous ligaments are a series of ligaments that course between the spinous processes of each pair of vertebrae from C2-3 to L4-5. Some authors consider this ligament to be the anterior aspect of the ligamentum nuchae in the cervical region (see following discussion). The interspinous ligaments are poorly developed in the cervical region, typically consisting of a thin, membranous, translucent septum (Panjabi et al., 1991). Thoracic interspinous ligaments are short from superior to inferior and broad from anterior to posterior and in the lumbar region the interspinous ligaments are more rectangular in shape (Standring et al., 2008). Because these ligaments are more fully developed in the thoracic region, they are discussed in more detail in Chapter 6.
Ligamentum Nuchae
The ligamentum nuchae (LN) is a flat, membranous structure that runs from the region between the cervical spinous processes anteriorly to the skin of the back of the neck posteriorly (Fig. 5-24), and spans the region between the occiput superiorly to the spinous process of C7 inferiorly. The posterior portion is its thickest and most distinct part and is sometimes called the funicular portion of the LN (or the dorsal nuchal raphe). This funicular part extends from the external occipital protuberance to the spinous process of C7. It is formed, at least in part, by the intertwining fibers of origin of the left and right trapezius, splenius capitis, and rhomboid minor muscles (Johnson, Zhang, & Jones, 2000; Mercer & Bogduk, 2003).
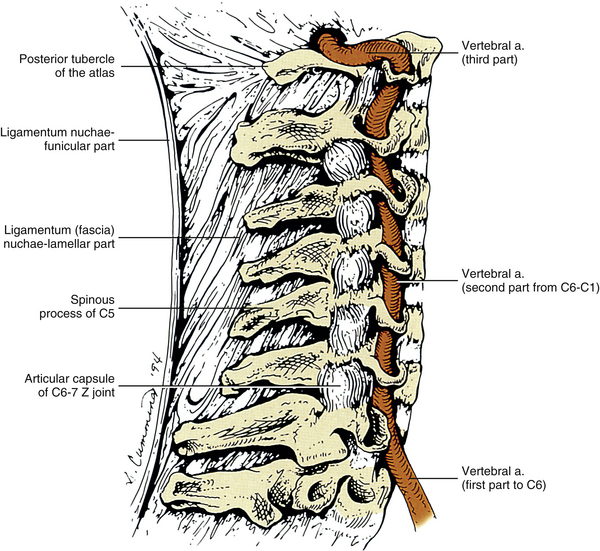
FIG. 5-24 Lateral view of the cervical portion of the vertebral column. The ligamentum (fascia) nuchae and the articular capsules of the C1-2 through C6-7 Z joints are seen. The vertebral artery can be seen entering the foramen of the transverse process (TP) of C6 and ascending through the remaining foramina of the TPs of C5 through C1. Then it can be seen passing around the superior articular process of C1. The vertebral artery disappears from view as it courses beneath the posterior atlanto-occipital membrane.
The thinner, larger, and more membranous anterior portion of the LN is known as the lamellar portion. The midline, fascial lamellar portion has two layers, left and right. The left and right layers attach anteriorly to the medial aspect of the left or right sides of the cervical bifid spinous processes, respectively. The small space between the two layers is filled with areolar (adipose) tissue (Kadri & Al-Mefty, 2007). Further anteriorly, the lamellar portion becomes continuous with the interspinous ligaments. Some authors consider the ligamentous tissue between the cervical spinous processes as supraspinous (along the posterior borders of adjacent spinous processes) and then interspinous ligaments (anteriorly between adjacent spinous processes) (Halliday et al., 1964), whereas others consider the lamellar portion of the LN to continue anteriorly to the base (anterior aspect) of the spinous processes, taking the place of the supraspinous and interspinous ligaments. The LN generally does take the place of the cervical supraspinous ligament, but the cervical interspinous ligaments are best thought of as distinct, albeit underdeveloped, cervical ligaments (Halliday et al., 1964). The lamellar portion of the LN also is continuous with the dense cervical fascia that extends laterally to separate the left and right semispinalis capitis muscles from the underlying semispinalis cervicis, multifidus cervicis, and suboccipital muscles. This dense cervical fascia extends superiorly and laterally to attach to the entire width of the occiput in the region of the inferior nuchal line (Mitchell, Humphreys, & O’Sullivan, 1998). Superiorly, the lamellar portion of the LN attaches to the occiput between the posterior aspect of the foramen magnum and the external occipital protuberance.
The LN is considered to be the homologue of the supraspinous and (by some authors) interspinous ligaments of the thoracic and lumbar regions and consequently limits flexion of the cervical region. In fact, the LN is considered “a significant structural restraint to flexion of the cervical spine” (Takeshita et al., 2004). The funicular portion of the LN also helps to maintain the cervical lordosis (Sakaura et al., 2008).
Between the occiput and C1 and between C1 and C2, the LN extends anteriorly all of the way to the dura mater, to which it attaches (Mitchell, Humphreys, & O’Sullivan, 1998; Dean & Mitchell, 2002; Humphreys et al., 2003). The attachment from the LN to the posterior dura mater between C1 and C2 can be seen on MRI (Humphreys et al., 2003). Figure 5-25 demonstrates the attachment between the LN and the posterior aspect of the spinal dura mater between C1 and C2 and its MRI appearance. These attachments may hold the dura mater posteriorly during cervical extension (to prevent buckling of the dura mater into the spinal cord) and flexion (to prevent the dura from moving forward and compressing the cord). Similar connective tissue attachments also connect the rectus capitis posterior minor muscle with the posterior aspect of the dura mater/posterior atlanto-occipital membrane complex (see Fig. 5-25). These latter attachments course between the occiput and the posterior arch of the atlas (see Chapter 4). Other connective tissue attachments to the posterior spinal dura mater have been identified and were described in the preceding section entitled Vertebral Canal.
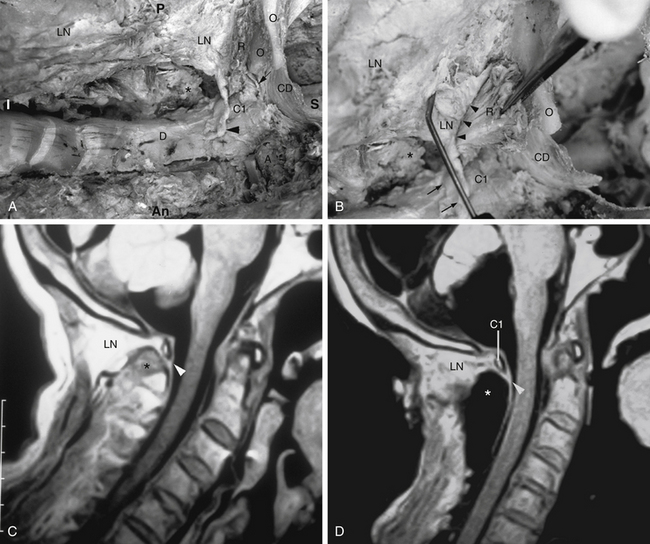
FIG. 5-25 A-D, Right lateral dissection of a cadaveric specimen. A, The spinous processes of the C2-6 vertebrae have been removed. The asterisk is located in the region normally occupied by the C2 spinous process. The attachment between the ligamentum (fascia) nuchae (LN) and (D) the dura mater of the cervical vertebral canal is identified with the large black arrowhead. The smaller black arrow demonstrates the attachment between (R), the rectus capitis posterior minor muscle (RCPM), and the superior-most aspect of the dura mater of the vertebral canal. A, vertebral artery; An, anterior; C1, posterior arch of the atlas; CD, dura mater within the posterior cranial fossa; I, inferior; O, occiput (note that a portion of the occiput has been sectioned in the midsagittal plane and the region labeled with a small “O” lies deep to and to the left of the plane of section); P, posterior; S, superior. B, Arrowheads demonstrate the attachment between the LN, being distracted posteriorly by the probe, and R, the posterior aspect of the RCPM being grasped by the forceps. This attachment is composed of a continuous series of firm connective tissue bridges coursing between the LN and RCPM. The small arrows and the region between the arrows at the bottom of this figure represent the attachment site of the LN to the dura mater. Pulling on the RCPM results in movement of the LN beginning in the region of the arrowheads. This movement continues to the region between the two arrows and results in tugging of the dura mater at this location. C, Magnetic resonance imaging (MRI) of the specimen in B before dissection was performed. Note the connection (arrowhead) between the LN and dura mater between the posterior arch of the atlas and spinous process of C2 (asterisk). D, MRI taken of this specimen after dissection. The spinous processes of the C2-6 vertebrae have been removed. The region normally occupied by the spinous process of C2 is indicated by an asterisk, and the posterior arch of the atlas is labeled C1. No contrast enhancement was used to produce this image. Notice that the LN retains the same appearance as in the predissection MRI of C as it passes between the posterior arch of the atlas and the region of the spinous process of C2. The connection of the LN with the dura mater (arrowhead) also retains the same appearance in the predissection and postdissection MRIs. By comparing this image with the same dissected specimen in A, one can observe that the LN is the only tissue inferior to Cl, the posterior arch of the atlas, as it passes inferior to the posterior arch of the atlas to attach to the dura mater. This helps confirm the MRI appearance of the LN in this region and, more specifically, the MRI appearance of the LN–dura mater attachment. (From Humphreys BK et al. [2003]. Investigation of connective tissue attachments to the cervical spinal dura matter. Clin Anat, 16, 152-159.)
In addition, between the occiput and the atlas the LN has a firm attachment along the entire posterior border of the rectus capitis posterior minor muscle (Humphreys et al., 2003).
Intertransverse Ligaments
Each intertransverse ligament passes from one transverse process to the transverse process of the vertebra below. These ligaments are not well-defined in the cervical region (Halliday et al., 1964) and frequently are replaced by the posterior intertransverse muscles. The thoracic intertransverse ligaments are rounded cords closely related to the deep back muscles (Standring et al., 2008). Some authors describe the lumbar intertransverse ligaments as being thin membranous bands. Others consider them to be discrete and well-defined. Still others consider them to consist of two distinct lamellae (Bogduk, 1997) (see Chapter 7).
General Considerations Related to the Role of Spinal Ligaments in Limiting Specific Motions
Notice from the preceding sections that in flexion, significant strains (tension) exist in all spinal ligaments, except the ALL. In extension, the ALL is under the most strain (the most stretched). In axial rotation the same-sided capsular ligament is maximally stretched (e.g., left axial rotation results in the highest strain being placed on the left capsular ligament). In lateral flexion, the contralateral ligamentum flavum and intertransverse ligament have the highest strains.
Flexion-Extension (Whiplash) Types of Injuries
Flexion-extension (whiplash) automobile injuries occur with some frequency. Even an 8-mile-per-hour rear end collision can cause injury to structures of the cervical region. Head rests are of limited value in preventing flexion-extension injuries, and in certain situations the head rest can actually act as a fulcrum and increase injury (Croft, 1993). In addition, the incidence of neck pain after flexion-extension injuries has been reported to be higher among those wearing seat belts (Lohiya, 1993). Although the brain is more susceptible to injury from side-to-side trauma (broadside accident) than from flexion-extension trauma (rear end collision), flexion-extension injuries can sprain the ligaments of the cervical region and lead to acute neck pain. In addition, flexion-extension injuries also can lead to chronic pain arising from the Z joints, temporomandibular joints, IVDs, and other structures of the neck (Croft, 1993; Barnsley et al., 1995; Lord et al., 1996). Croft (1993) reports that 45% to 83% of patients involved in flexion-extension injuries have symptoms that persist for more than 2 years after litigation related to their case has been settled (i.e., these symptoms are not related to pending litigation). Disc pathology seems to contribute to chronic neck pain after flexion-extension (whiplash) injuries (Pettersson et al., 1997), and Loudon and colleagues (1997) found evidence of a decrease in proprioceptive ability after flexion-extension injuries. They speculate that this results from damage to cervical neck muscles and to the Z joint capsules, resulting in abnormal proprioception from these structures.