Bone Marrow, Blood Cells, and the Lymphatic System*
Bone Marrow and Blood Cells
In adults, the bone marrow is the primary site of blood cell production, or hematopoiesis, which is a term is derived from haima (Gr., blood) and poiein (Gr., to make). Hematopoiesis may also occur elsewhere (extramedullary hematopoiesis), most commonly in the spleen. Hematopoiesis is also known as hemopoiesis.
Hematopoiesis is first seen as blood islands in the embryonic yolk sac. During gestation, the main site of hematopoiesis shifts from the yolk sac to the liver and spleen and eventually to the bone marrow. The composition of the marrow changes with age. The general pattern is that hematopoietic tissue (red marrow) regresses and is replaced with nonhematopoietic tissue, mainly fat (yellow marrow). Thus, in newborns and very young animals, the bone marrow consists largely of hematopoietically active tissue, with relatively little fat, whereas in geriatric individuals the marrow consists largely of fat. In adults, hematopoiesis occurs primarily in the pelvis, sternum, ribs, vertebrae, and the proximal ends of humeri and femora. Even within these areas of active hematopoiesis, fat may constitute a significant proportion of the marrow volume.
The marrow cavity is interlaced with venous sinuses composed of a luminal layer of specialized endothelial cells and an abluminal layer of specialized fibroblasts known as adventitial reticular cells (Figs. 13-1 and 13-2). Hematopoiesis takes place between these venous sinuses in the so-called hematopoietic spaces. Sinusoidal endothelial cells function as a barrier and regulate traffic of chemicals and particles between the intravascular and extravascular spaces. Reticular cells extend cytoplasmic processes into the hematopoietic spaces, forming a scaffold that supports the hematopoietic cells. Osteoblasts, osteoclasts, and elongated flat cells line the endosteum. Stromal cells, presumably of fibroblast origin, provide structural support and produce extracellular matrix components such as proteoglycans, glycosaminoglycans, collagen, fibronectin, and others. Adipocytes, macrophages, lymphocytes, and plasma cells are also components of the hematopoietic microenvironment.
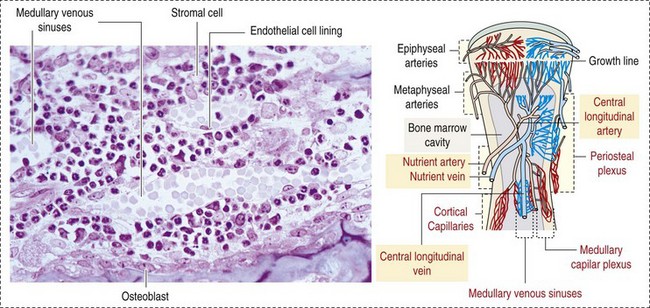
Fig. 13-1 Bone marrow: Structure and vascularization. (From Kierszenbaum AL: Histology and cell biology: an introduction to pathology, ed 28, St Louis, 2007, Mosby.)
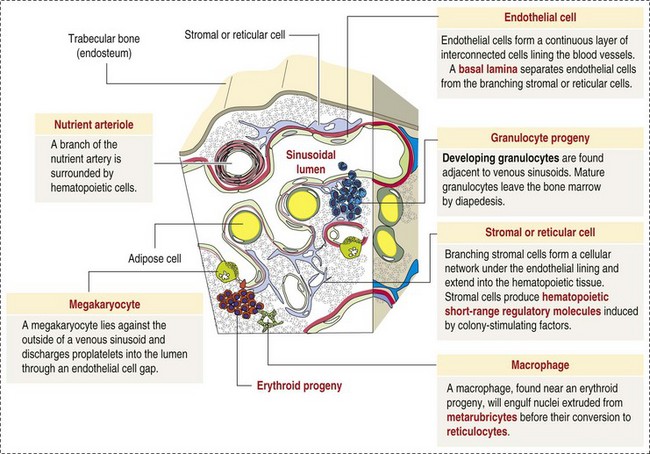
Fig. 13-2 Bone marrow: Structure. (Adapted from Kierszenbaum AL: Histology and cell biology: an introduction to pathology, ed 2, St Louis, 2007, Mosby.)
Hematopoietic cells, supporting connective tissue cells, extracellular matrix components, and soluble factors form the bone marrow hematopoietic microenvironment. There is complex functional interplay among these components. Behavior of hematopoietic cells is influenced by direct cell-to-cell and cell-matrix interactions and by soluble mediators, such as cytokines and hormones, that interact with cells and with matrix proteins. Cells localize to specific niches within the hematopoietic microenvironment via adhesion molecules, such as integrins, immunoglobulins (Igs), lectins, and other receptors, that recognize ligands on other cells or matrix components. Cells also express receptors for soluble molecules such as chemokines (chemoattractant cytokines) and hormones that influence cell trafficking and metabolism.
The following basic concepts provide a framework for understanding the mechanisms of injury and diseases presented later in the chapter.
• Hematopoietic tissue is highly proliferative. Billions of cells per kilogram of body weight are produced each day.
• Pluripotent hematopoietic stem cells are a self-renewing population, giving rise to cells with committed differentiation programs, and are common ancestors of all blood cells. The process of hematopoietic differentiation is shown in Fig. 13-3.
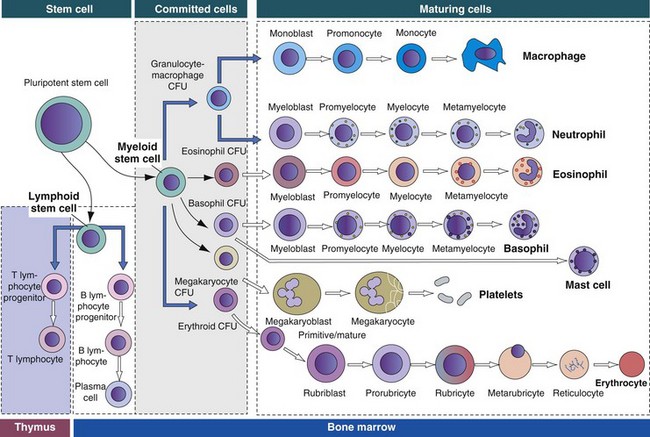
Fig. 13-3 Schematic illustration of the hematopoietic hierarchy in the bone marrow.
The bone marrow consists of (1) stem cells, pluripotent cells capable of self-renewal; (2) committed progenitor cells (myeloid and lymphoid progenitor cells); and (3) maturing cells. Maturing cells develop from cells called colony-forming units (CFUs). The myeloid stem cell gives rise to CFUs responsible for the regeneration of red blood cells (erythroid CFUs), platelets (megakaryocyte CFUs), basophils (basophil CFUs), and eosinophils (eosinophil CFUs). Monocytes and neutrophils are derived from a common committed progenitor cell (granulocyte-macrophage CFU). The lymphoid progenitor cell generates the B lymphocyte progeny in the bone marrow and T lymphocyte progeny in the thymus. (Adapted from Kierszenbaum AL: Histology and cell biology: an introduction to pathology, St Louis, 2002, Mosby.)
• Hematopoietic cells undergo sequential divisions as they develop, so there are progressively higher numbers of cells as they mature. Cells also continue to mature after they have stopped dividing. Conceptually, it is helpful to consider cells in the bone marrow as belonging to mitotic and postmitotic compartments. Examples of developing hematopoietic cells are shown in Fig. 13-4.
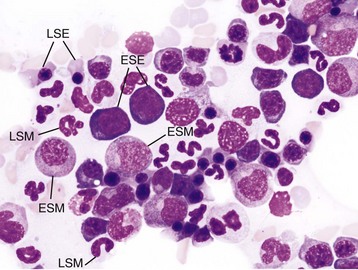
Fig. 13-4 Normal hematopoiesis, canine bone marrow aspirate.
Wright’s stain. ESE, Early stage erythroid; ESM, early stage myeloid; LSE, late stage erythroid; LSM, late stage myeloid. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
• Mature cells released into the blood circulation have different normal lifespans, varying from hours (neutrophils), to days (platelets), to months (erythrocytes), and to years (some lymphocytes).
• The hematopoietic system is under exquisite local and systemic control and responds rapidly and predictably to various stimuli.
• Production and turnover of blood cells are balanced so that numbers are maintained within normal ranges (steady-state kinetics) in healthy individuals.
• Normally the bone marrow releases only mature cell types (and very low numbers of cells that are almost fully mature) into the circulation. In response to certain physiologic or pathologic stimuli, however, the bone marrow releases immature cells that are further back in the supply “pipeline.”
Regulation of Hematopoiesis
Control of hematopoiesis is complex, with many redundancies, feedback mechanisms, and pathways that overlap with other physiologic and pathologic processes. Many cytokines influence cells of different lineages and stages of differentiation. The aim of this section is not to explain these regulatory pathways in detail but rather to provide a broad overview and thus a basic framework for understanding mechanisms of disease involving the hematopoietic system.
Erythropoiesis
Erythropoiesis—from erythros (Gr., red)—refers to the production of red blood cells (RBCs), or erythrocytes, whose main function is gas (oxygen [O2] and carbon dioxide [CO2]) exchange. The dominant regulator of erythropoiesis is a glycoprotein aptly named erythropoietin (Epo). Epo acts on early erythroid progenitor cells in concert with other cytokines, including interleukins (IL-3, IL-4, and IL-9), granulocyte-macrophage colony-stimulating factor (GM-CSF), and insulin-like growth factor. Epo is synthesized primarily in the kidney, and exerts its effects by promoting proliferation and inhibiting apoptosis of developing erythroid cells that express Epo receptors. The stimulus for increased Epo production is hypoxia. Because erythrocytes are packed with hemoglobin, of which iron is an essential component, erythropoiesis requires adequate availability of iron. Physiologic systems exist to conserve and recycle iron, as discussed further later.
The earliest erythroid precursor identifiable by routine light microscopy is the rubriblast, which undergoes maturational division to produce 8 to 32 progeny cells. Late-stage erythroid precursors known as metarubricytes extrude their nuclei and become reticulocytes, which are the cells at the differentiation stage immediately preceding the mature erythrocyte. Reticulocytes start maturing in the bone marrow and finish maturing in the peripheral blood circulation and the spleen. (Horses are an exception in that they do not release reticulocytes into circulation, even in situations of increased demand.) The normal transit time from rubriblast to mature erythrocyte is approximately 1 week. Unlike mature erythrocytes, which lack organelles, reticulocytes still contain ribosomes and mitochondria, mainly to support completion of hemoglobin synthesis. These remaining organelles impart a bluish-purple cast (polychromasia) to reticulocytes on routine blood smear examination and when stained with dye, such as new methylene blue, precipitate to form irregular aggregates of dark-staining material (Fig. 13-5). Cats also have a more mature form of reticulocyte in circulation, the punctate reticulocyte, which has a much finer staining pattern when stained with new methylene blue and does not appear bluish-purple (polychromatophilic) on routine blood smear examination (see the section Anemia).
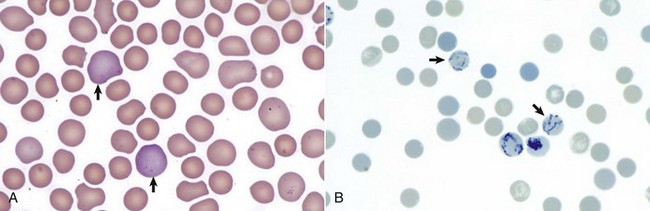
Fig. 13-5 Reticulocytosis, canine blood smears.
A, Reticulocytes (arrows) appear polychromatophilic with routine staining. Wright’s stain. B, Reticulocytes. Precipitated aggregates of RNA are stained blue (arrows) with new methylene blue. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Mature erythrocytes circulate for a long time compared with other blood cells. The erythrocyte mean lifespan varies between species: approximately 150 days in horses and cattle, 100 days in dogs, and 70 days in cats. Erythrocytes therefore must be highly resilient cells capable of enduring continual mechanical and biochemical stresses. In most mammals, erythrocytes have a biconcave disk shape, sometimes called the discocyte. Species in which the discocyte is not the norm include goats (flat, irregularly shaped RBCs), camelids (oval RBCs), and some cervids (sickle-shaped RBCs). In dogs, some breeds (Akitas and Shibas) have smaller erythrocytes than others; erythrocytes in these breeds also have a high concentration of potassium, unlike erythrocytes in other dogs.
One of the key properties of erythrocytes is deformability; they change shape as they move through the microvasculature. This deformability is a function of interactions between the cell membrane, the cytoskeleton, and intracellular contents. Mature mammalian erythrocytes lack nuclei and organelles and are thus incapable of transcription, translation, and oxidative metabolism. However, they do require energy for various functions, including maintenance of shape and deformability, active transport, and prevention of oxidative damage, and they generate this energy entirely through glycolysis (also known as the Embden-Meyerhof pathway). Erythrocyte antioxidant biochemical pathways are discussed in more detail in the section on Defense Mechanisms.
The concentration of circulating erythrocytes typically decreases postnatally and remains below normal adult levels during the period of rapid body growth. The age at which erythrocyte numbers begin to increase and the age at which adult levels are reached vary among species. In dogs, adult values are usually reached between 4 and 6 months of age; in horses, this occurs at approximately 1 year of age. In most species, erythrocytes are larger at birth, and their mean volume decreases as fetal erythrocytes are replaced.
Granulopoiesis and Monocytopoiesis (Myelopoiesis)
Granulocytes (neutrophils, eosinophils, and basophils) and monocytes have key immunologic functions, including phagocytosis and microbicidal activity (neutrophils and monocyte-derived macrophages); parasiticidal activity and participation in allergic reactions (eosinophils and basophils); antigen processing and presentation; and cytokine production (macrophages). Inflammatory mediators, such as ILs and tumor necrosis factor α (TNF-α), stimulate fibroblasts, macrophages, and endothelial cells to produce cytokines, such as granulocyte colony-stimulating factor (G-CSF) and GM-CSF, that increase granulopoiesis and monocytopoiesis.
Granulocytic and monocytic cells are sometimes referred to collectively as myeloid cells. (note: This terminology can be confusing because “myeloid” also has other meanings. In the most general sense, it refers to the bone marrow. In the context of hematology, “myeloid” is also sometimes used to mean a hematopoietic cell of nonlymphoid origin, a distinction reiterated in the classification of leukemias as discussed later in this chapter. In neuroanatomy, the prefixes “myelo-” or “myel-” can refer to the spinal cord or myelin.) The earliest granulocytic or monocytic precursor identifiable by routine light microscopy is the myeloblast, which undergoes maturational division to produce 16 to 32 progeny cells. The normal transit time from myeloblast to mature neutrophil is approximately 5 days. In addition, a reserve of fully mature neutrophils is maintained in the bone marrow. The size of this so-called storage pool is species-dependent. (Species differences and clinical significance of the storage pool are discussed in more detail in later sections.)
The concentration of granulocytes measured in the blood depends on the rate of production and release from the bone marrow, the proportions of cells circulating freely within the vasculature versus those transiently attached to the endothelial surface (marginated), and the rate of migration from the vasculature into tissues. Neutrophils normally are the predominant leukocyte type in blood of most domestic species. Neutrophils are only in circulation for a short time (less than 12 hours).
Thrombopoiesis
Thrombopoiesis—from thrombos (Gr., clot)—refers to the production of platelets, which have a central role in primary hemostasis and also participate in coagulation and inflammatory pathways. Thrombopoietin (Tpo), synthesized primarily in the liver, is the dominant regulator of thrombopoiesis. Unlike Epo, which is upregulated in response to hypoxia, Tpo is produced at a relatively constant rate (constitutively). The mechanism regulating production is therefore different. Platelets and their precursors express the Tpo receptor, which binds Tpo in the plasma. When the platelet mass is decreased, less Tpo is bound to platelet receptors, and the increased concentration of Tpo free in the plasma stimulates thrombopoiesis.
Thrombopoiesis differs fundamentally from other forms of hematopoiesis. Platelets (sometimes called thrombocytes) are anucleate cells formed not by maturational division but by shedding of membrane-bound cytoplasmic fragments from precursor cells called megakaryocytes. (note: Birds and reptiles produce thrombocytes in the manner of other blood cells and their thrombocytes and erythrocytes are nucleated.) As the name suggests, megakaryocytes are very large cells, much larger than any other hematopoietic cells (Fig. 13-6). Megakaryocytes arise from a progenitor cell and undergo endomitosis to become polyploid (usually 8N-32N). As they mature, megakaryocytes increase in size and their cytoplasm becomes more abundant. They change in staining pattern (Wright’s stain of aspirates) from intensely basophilic to eosinophilic, and their nuclei change from round to highly lobulated. Megakaryocytes extend cytoplasmic processes into the lumina of the bone marrow venous sinusoids, where they shed platelets into circulation. The lifespan of a platelet in circulation is approximately 6 days in the dog.
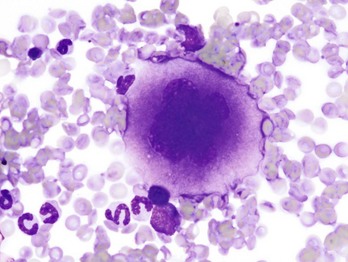
Fig. 13-6 Megakaryocyte, canine bone marrow aspirate.
Note the cell’s very large size, lobulated nucleus, and abundant granular cytoplasm. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Platelets normally circulate in a quiescent state, as flattened disks. Activation of platelets is triggered by soluble agonists, such as thrombin, or insoluble agonists, such as collagen. Platelets express surface receptors for fibrinogen, von Willebrand factor (vWF), collagen, and other ligands. Other key features of platelets include a network of membrane invaginations known as the open canalicular system (not present in cattle), which facilitates the expansion of surface area and release of granule contents in response to activation; specialized endoplasmic reticulum known as the dense tubular system that acts as a reservoir for calcium; and cytoskeletal components involved in mediating shape change, release of granule contents, receptor interactions, and clot retraction. When activated, platelets change to more spherical forms with pseudopodia and participate in a number of related processes at sites of vascular damage:
• Adhesion to exposed subendothelial collagen
• Secretion of bioactive granule contents (including calcium, adenosine diphosphate [ADP], fibrinogen, vWF, histamine, serotonin, and others)
• Aggregation with other platelets
• Eventual fusion with other platelets to form a platelet plug
Phospholipids (especially phosphatidylserine) expressed on the surface of activated platelets also help to localize coagulation (formation of an insoluble fibrin clot) to sites of vascular damage.
Lymphopoiesis
Lymphopoiesis refers to the production of new lymphocytes. T lymphocytes and B lymphocytes are the key effectors of cell-mediated and humoral immunity, respectively. Lymphopoiesis is discussed in more detail in Chapter 5, but a brief overview is provided here. T lymphocytes originate in the bone marrow and migrate to the thymus, where they undergo differentiation, selection, and maturation processes before migrating to the peripheral lymphoid tissue as effector cells. B lymphocyte development occurs in two phases, first in an antigen-independent phase in the bone marrow and ileal Peyer’s patches (the site of B lymphocyte development in ruminants), then in an antigen-dependent phase in peripheral lymphoid tissues (such as spleen, lymph nodes, and mucosa-associated lymphoid tissue [MALT]). Trafficking of lymphocytes occurs under the direction of chemokine (chemoattractant cytokine) signals. Once they have migrated to the peripheral lymphoid tissue, lymphocytes may undergo clonal expansion in response to antigenic stimulation.
Unlike other hematopoietic cells, which circulate only in the blood vessels, lymphocytes travel in both blood and lymphatic vessels, continually recirculating between the two systems. In most species, the majority of lymphocytes in blood circulation are T lymphocytes. According to conventional wisdom, cattle normally have higher numbers of lymphocytes than neutrophils in circulation. However, recent work suggests that is no longer the case, at least in mid-lactation Holstein cows, most likely because of changes in genetics and husbandry.
Hemostasis (Coagulation And Platelet Function)
Hemostasis—from haima (Gr., blood) and stasis (Gr., “a standing still”)—refers to the cessation of bleeding. The main components responsible for hemostasis are platelets, blood vessels, and the coagulation system. Bleeding disorders may result if any of these systems are perturbed. Although coagulation factors are mainly synthesized in the liver, they are included in this chapter because of their inextricable link with the hematopoietic system.
Physiologic hemostasis involves coordinated processes: formation of a platelet plug at the site of vascular damage (primary hemostasis), development of a cross-linked network of insoluble fibrin that provides further clot stability (secondary hemostasis) and retraction and enzymatic degradation of the clot (tertiary hemostasis). The main components of these processes are summarized in Chapter 2 and Figs. 2-12 through 2-17. Signalment, history, and clinical signs are important in evaluating any patient with a suspected bleeding tendency, but laboratory testing is required to definitively identify specific defects in hemostasis. Routine assays for assessing hemostasis are summarized in Web Appendix 13-1.
Blood Vessels: Blood vessels have numerous functions in hemostasis. Endothelial cells produce and metabolize many molecules involved in promoting and regulating hemostasis (see Fig. 2-16). For example, they produce prothrombotic molecules, such as vWF and thromboxane A2; antithrombotic molecules, such as prostacyclin (PGI2), thrombomodulin, and nitric oxide; and fibrinolytic molecules, such as tissue plasminogen activator. When a blood vessel is damaged, a rapid, transient neuromechanical vasoconstriction occurs in response. Collagen underlying the damaged endothelium provides structural support for platelet adhesion and clot formation, and also acts as a key biochemical activator of platelets and the intrinsic pathway of the coagulation system (see next section).
Coagulation System: Stopping hemorrhage from a damaged blood vessel ultimately depends on the formation of a stable fibrin clot. This process depends on a number of plasma proteins, or clotting factors, that normally circulate in an inactive proenzyme state. These clotting factors, together with other molecules, including nonenzymatic proteins, calcium, and platelet phospholipids, are known collectively as the coagulation system. Activation of the coagulation system initiates a series of linked enzymatic reactions that result in the production of thrombin. Thrombin converts the soluble plasma protein fibrinogen to fibrin, which in turn is enzymatically cross-linked to form an insoluble clot. The coagulation system is conventionally considered to consist of three interrelated pathways for ease of understanding and interpretation of test results. These pathways—the intrinsic, extrinsic, and common pathways—are summarized in Fig. 2-13.
Anticoagulant and fibrinolytic pathways provide critical “checks and balances.” These pathways are initiated simultaneously with activation of the coagulation system. Procoagulant and anticoagulant pathways are regulated at multiple levels and normally maintained in a balance that avoids hypercoagulable or hypocoagulable states. Some key regulators of hemostasis are listed in Web Box 13-1. Counterregulatory mechanisms are critical for the prevention of inappropriate or excessive clotting and the eventual dissolution of the clot (fibrinolysis) after it has formed. Fibrinolysis results from the action of plasmin on fibrin (see Figs. 2-14, 2-16, and 2-17). The inactive proenzyme plasminogen is converted to plasmin by several molecules, including tissue plasminogen activator, factor XII, and thrombin. Note that some of these molecules (e.g., factor XII and thrombin) have both procoagulant and anticoagulant effects. There are also regulators of fibrinolysis, including inhibitors of plasminogen and plasmin. Fibrinolysis results in production of fibrin degradation products (FDPs). D-dimer is a specific type of FDP that results from the degradation of insoluble cross-linked fibrin. Fibrinolytic products (FDPs and D-dimer) in the blood are often measured to detect inappropriate coagulation and are also of clinical importance because they can impair hemostasis by inhibiting fibrin polymerization and platelet function.
Platelets: The basics of platelet function are described in the previous section on Thrombopoiesis. Platelets normally circulate in a quiescent state, as flattened disks. Activation of platelets is triggered by soluble agonists, such as thrombin, or insoluble agonists, such as collagen. Platelets express surface receptors for fibrinogen, vWF, collagen, and other ligands. Other key features of platelets include a network of membrane invaginations known as the open canalicular system (not present in cattle), which facilitates the expansion of surface area and release of granule contents in response to activation; specialized endoplasmic reticulum known as the dense tubular system that acts as a reservoir for calcium; and cytoskeletal components involved in mediating shape change, release of granule contents, receptor interactions, and clot retraction. When activated, platelets change to more spherical forms with pseudopodia and participate in a number of related processes at sites of vascular damage:
• Adhesion to exposed subendothelial collagen
• Secretion of bioactive granule contents (including calcium, adenosine diphosphate [ADP], fibrinogen, vWF, histamine, serotonin, and others)
• Aggregation with other platelets
• Eventual fusion with other platelets (a process known as viscous metamorphosis) to form a platelet plug
Phospholipids (e.g., phosphatidylserine) expressed on the surface of activated platelets also help to localize the clot to sites of vascular damage.
Portals of Entry
Invading cells or microorganism gain access to the bone marrow or blood circulation either hematogenously or by trauma. Trauma may be as obvious as a gaping wound or as subtle as the bite of an insect. Portals of entry for the bone marrow are summarized in Box 13-1.
Defense Mechanisms
The bone marrow is encased by a protective shell of cortical bone, and blood supply to the marrow provides access to systemic humoral and cellular defenses. Of course, leukocytes themselves function as an essential part of inflammation and immune function, as discussed briefly in the Granulopoiesis and Monocytopoiesis section and in greater detail in Chapters 3 and 5.
Biochemical steps in the glycolytic pathway or linked to it generate antioxidant molecules that enable erythrocytes to withstand oxidative insults throughout their many days in circulation. In addition to producing energy in the form of adenosine triphosphate (ATP), glycolysis generates reduced nicotinamide adenine dinucleotide (NADH), which helps convert the oxidized, nonfunctional form of hemoglobin, known as methemoglobin, back to its active reduced state. Another antioxidant erythrocyte metabolic pathway, the pentose shunt or hexose monophosphate shunt pathway, generates reduced nicotinamide adenine dinucleotide phosphate (NADPH) to help maintain glutathione in the reduced state.
Responses to Injury
The main mechanisms of disease in the bone marrow and blood cells are shown in Box 13-2, and are discussed in more detail here.
Bone Marrow
Bone marrow responses to injury include abnormal patterns of proliferation of normal hematopoietic cell types, proliferations of abnormal hematopoietic cell types (neoplastic or nonneoplastic), replacement of normal hematopoietic tissue by abnormal cells or fibrous tissue, necrosis, and inflammation. Each of these is discussed further; the relationship between altered hematopoiesis and concentrations of blood cells in circulation is discussed in more detail in the next section on Blood Cells.
The classic nomenclature for describing different types of abnormal cellular proliferations applies to the hematopoietic system, with some exceptions and modifications. Hyperplasia, an increase in cell number, may be an obviously appropriate response to a stimulus (e.g., erythroid hyperplasia in response to hypoxemia, or granulocytic hyperplasia in response to inflammatory stimuli). In other situations, hyperplasia is a secondary phenomenon for which the specific stimulus may not be clear—for example, a number of conditions (inflammation, neoplasia, iron deficiency, asplenia, and others) are associated with reactive thrombocytosis, in which an increased concentration of circulating platelets reflects megakaryocytic hyperplasia. Sometimes hyperplasia is idiopathic. For example, splenic nodular hyperplasia, which often includes a hematopoietic component, is frequently an incidental finding.
Hypertrophy, an increase in cell size, is not a conventional description for hematopoietic cells or for bone marrow as an organ. Abnormally large hematopoietic cells are considered evidence of dysplasia, or altered cell formation (alterations to shape or organization of cells are also evidence of dysplasia). Dysplasia of hematopoietic cells, indicating dyshematopoiesis, may be primary (idiopathic) or secondary to a number of conditions, including infection, nutritional imbalance, and toxicosis. Unfortunately, the terminology is sometimes confusing. For example, myelodysplastic syndromes are clonal hematopoietic stem cell disorders that are in fact neoplastic (see the section on Hematopoietic Neoplasia). Splenic hypertrophy is usually referred to as splenomegaly and may reflect processes other than hyperplasia or cellular hypertrophy (e.g., congestion). Similarly, lymph node enlargement is typically referred to as lymphadenopathy or lymphadenomegaly and may reflect lymphoid hyperplasia or other processes (e.g., neoplasia, inflammation, or edema).
Replacement of hematopoietic tissue in the bone marrow by abnormal tissue, usually fibrous tissue or malignant cells, is known as myelophthisis. The term metaplasia, describing the replacement of cells normally comprising a tissue with another well-differentiated cell type, is rarely used to describe hematopoietic tissues in veterinary medicine, although in humans, the terms myeloid metaplasia and extramedullary hematopoiesis are sometimes used interchangeably. Similarly, the term atrophy, which means a diminution in the size of a cell, tissue, organ, or part, is seldom used to describe bone marrow. Instead, a decrease in bone marrow hematopoietic tissue is typically referred to as hypoplasia, as discussed later in the section on cytopenias. The absence of bone marrow hematopoietic tissue of a particular lineage is typically referred to as aplasia or, if affecting all lineages, by the misleading term aplastic anemia (more aptly termed aplastic pancytopenia). An exception is “serous atrophy of fat,” a condition associated with cachexia or starvation in which fat is catabolized and bone marrow reticular cells produce a mucoid ground substance (Web Fig. 13-1). However, neither hematopoietic cells nor fat is necessarily absent from the bone marrow in this condition, and other names, such as gelatinous transformation, have been suggested to describe it. Neoplasia of bone marrow or lymphoid organs may be primary or secondary (metastatic). Neoplasia may arise from any hematopoietic lineage. Specific types of hematopoietic neoplasia (leukemias, lymphomas, and others) are covered later in this chapter.
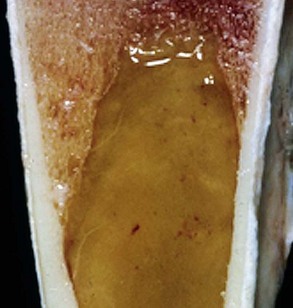
Web Fig. 13-1 Serous atrophy of fat (gelatinous transformation), bone marrow, calf.
Serous atrophy of fat is caused by starvation and/or cachexia and is characterized by replacement of normal fat and hematopoietic elements by a gelatinous extracellular matrix. (Courtesy Department of Pathobiology, College of Veterinary Medicine, University of Tennessee).
Bone marrow fibrosis (myelofibrosis) and necrosis are associated with many common underlying conditions, often occur in tandem, and are described most extensively in dogs. Conditions known or suspected to cause myelofibrosis and necrosis include sepsis, malignancy (especially hematopoietic malignancies), drug toxicity, and immune-mediated disease (especially nonregenerative immune-mediated hemolytic anemia [IMHA]).
Bone marrow inflammation can take different forms. Granulomatous inflammation in response to fungal infection (e.g., histoplasmosis) or mycobacteriosis may be evident histologically or as macrophage hyperplasia in cytologic preparations of aspirates. Phagocytosed organisms within macrophages are detectable with either type of sample. Dogs and cats with nonregenerative IMHA often have bone marrow inflammation, in addition to fibrosis and necrosis. The inflammation is evident as fibrin deposition, edema, and multifocal neutrophilic infiltrates; immune-mediated cytopenias are also associated with bone marrow lymphocytic and/or plasma cell hyperplasia.
Blood Cells
Responses of circulating blood cells to injury include decreased survival (destruction, consumption, or loss), altered distribution, and altered structure or function (or both). These responses are not mutually exclusive—for example, altered erythrocyte structure may lead to decreased survival. Often, but not always, these responses result in decreased concentrations of blood cells in circulation.
Abnormal Concentrations of Blood Cells: Cytopenia—from kytos (Gr., hollow vessel) and penia (Gr., poverty)—refers to a deficiency of blood cells in circulation, hence the terms neutropenia, thrombocytopenia, lymphopenia, and so on. Basic mechanisms causing cytopenias include decreased production (hypoplasia), increased destruction, blood loss, consumption, and altered anatomic distribution (e.g., shifts between marginated and circulating compartments in the blood, or between the spleen and the peripheral blood circulation). Most of these basic mechanisms may cause cytopenias of more than one lineage, and some processes affect multiple lineages simultaneously. Sustained decreased production of all three major bone marrow hematopoietic lineages results in pancytopenia (anemia, neutropenia, and thrombocytopenia). Pancytopenia may occur because of myelophthisis, in which normal bone marrow tissue is replaced by abnormal cells or tissue (as in the case of effacement by malignant cells or fibrous tissue) or because of an abnormality of hematopoietic cells themselves. Destruction of hematopoietic stem cells and progenitor cells causes a condition known as aplastic anemia, or aplastic pancytopenia, which is discussed further in the section on specific diseases.
The pattern of development of cytopenias is partly a function of normal blood cell kinetics. The lifespan of different blood cells in circulation varies markedly (neutrophils: hours, platelets: days, erythrocytes: months). Thus severe neutropenia typically develops within 1 week after cessation of granulopoiesis and after the bone marrow storage pool of neutrophils is depleted, whereas severe thrombocytopenia typically develops in the second week after cessation of thrombopoiesis. Anemia develops much more slowly after the cessation of erythropoiesis, if at all, depending in part on how rapidly the marrow recovers from the insult and on species variation in the erythrocyte lifespan.
Increases in the concentration of many blood cells are indicated by the suffix, -osis (erythrocytosis, lymphocytosis, monocytosis, and thrombocytosis). Increases in granulocytes are indicated by the suffix, -philia (neutrophilia, eosinophilia, and basophilia). Basic mechanisms causing increases in the concentration of blood cells vary considerably according to cell type and are discussed in more detail later. Neoplasia can result in increased or decreased numbers of blood cells. In many kinds of hematopoietic neoplasia, especially leukemias, there are readily detectable numbers of neoplastic cells in the blood. Specific types of hematopoietic neoplasia are discussed later in this chapter. Increased concentrations of blood cells can also occur secondary to many forms of neoplasia (i.e., as a paraneoplastic syndrome), including non-hematopoietic neoplasms, because of production of stimulatory cytokines by the tumor.
Veterinary laboratories typically provide species-specific hematology reference values based on a reference population of clinically normal adult animals. However, it is important to note that what is normal may vary not only between species but also as a function of other factors, such as age, breed, geographic location, and differences in laboratory methodology.
Mechanisms of disease causing cytopenias and abnormally increased concentrations of specific types blood cells are covered in more detail in the next section.
Anemia: Anemia refers to subnormal red blood cell mass or hemoglobin concentration. Anemia causes clinical signs referable to decreased oxygen-carrying capacity (pallor of mucous membranes, lethargy, weakness, exercise intolerance) and may also result in detectable abnormalities because of tissue hypoxia (e.g., increased liver enzyme activities as a result of hypoxia-induced damage to hepatocytes). Anemia also causes decreased viscosity of the blood, and in marked cases frequently causes heart murmurs as a result of a decrease in laminar blood flow.
Classifying anemia as regenerative or nonregenerative is clinically useful because it provides information about the mechanism of disease (Table 13-1). The hallmark of regenerative anemias, except in horses, is reticulocytosis (i.e., increased numbers of circulating reticulocytes [immature erythrocytes]), which is evident as polychromasia on a routinely stained blood smear. In ruminants, reticulocytosis is often accompanied by basophilic stippling (Fig. 13-7).
TABLE 13-1
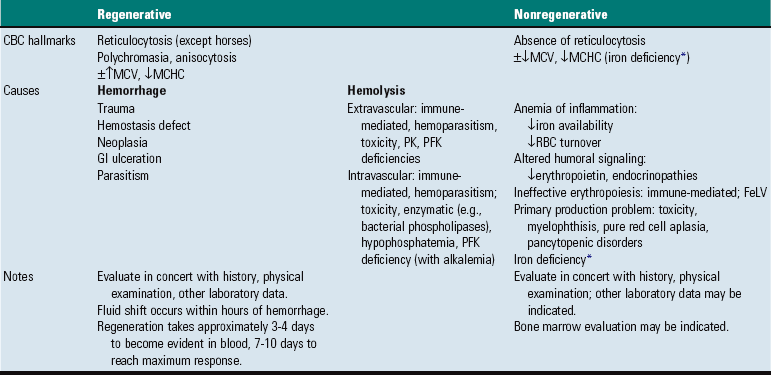
CBC, Complete blood count; FeLV, feline leukemia virus; GI, gastrointestinal; MCHC, mean cell hemoglobin concentration; MCV, mean cell volume; PFK, phosphofructokinase; PK, pyruvate kinase; RBC, red blood cell.
*Iron deficiency anemia is sometimes regenerative
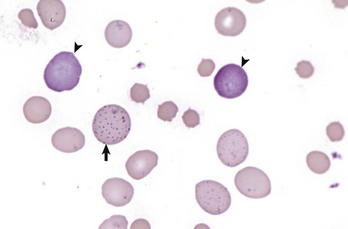
Fig. 13-7 Basophilic stippling and polychromasia, bovine blood smear.
Erythrocytes from this cow with regenerative anemia include several cells with basophilic stippling (arrow) and two polychromatophilic cells (reticulocytes) (arrowheads). Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Reticulocytosis indicates increased bone marrow erythropoiesis (Fig. 13-8) and release of erythrocytes before they are fully mature (further back in the production “pipeline”). Reticulocytosis is an appropriate response to anemia. A strong regenerative response may produce an increased mean cell volume (MCV) and decreased mean cell hemoglobin concentration (MCHC) on the complete blood count (CBC), because reticulocytes are larger and have a lower hemoglobin concentration than mature erythrocytes. Horses are an exception to this classification scheme because they do not release reticulocytes into circulation even when their marrow is producing increased numbers of erythrocytes. Horses with a regenerative response may have an increased MCV and red cell distribution width (an index of variation in cell size). But definitive determination of regeneration in a horse requires either bone marrow examination, in which case the evidence of regeneration is erythroid hyperplasia, or sequential CBCs, in which case the evidence of regeneration is an increase in red cell mass over time. Punctate reticulocytes in cats are evident when blood is stained with new methylene blue, but they are not counted as reticulocytes on the CBC and have the same appearance as mature erythrocytes on routine blood smear examination.
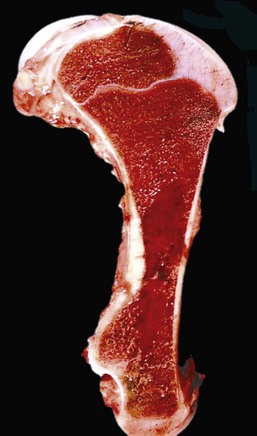
Fig. 13-8 Hemopoietically active bone marrow, femur, calf.
Note that the bone marrow has a uniform consistency and is red to dark red. These responses are characteristic of hemopoietically active bone marrow. (Courtesy Dr. Ramos, Autonomous University of Barcelona; and Noah’s Arkive, College of Veterinary Medicine, University of Georgia.)
Another finding that may accompany regeneration, in addition to reticulocytosis, is the presence of nucleated erythroid cells (nRBCs). However, the presence of circulating nRBCs is not in itself definitive evidence of regeneration and in fact may signify dyserythropoiesis (e.g., because of lead poisoning or bone marrow disease) or splenic dysfunction. When nRBCs are present as part of a regenerative response to anemia, they should be in low numbers relative to the numbers of reticulocytes.
Recall that the stimulus for increased erythropoiesis is increased secretion of Epo in response to hypoxemia. Although the action of Epo on erythropoiesis is rapid, evidence of a regenerative response is not immediately apparent in a blood sample. One of the main effects of Epo is to expand the pool of early stage erythroid precursors, and it takes time for these cells to differentiate to the point where they are released into circulation. In a case of acute blood loss, for example, it typically takes 3 to 4 days until reticulocytosis is evident on the CBC, and several more days until the regenerative response peaks. A similar lag occurs in cases of acute hemolysis. The term preregenerative is sometimes used to describe anemia with a regenerative response that is impending but not yet apparent on the CBC. Confirming a regenerative response in such cases requires either evidence of erythroid hyperplasia in the bone marrow or emergence of a reticulocytosis on subsequent days.
Regenerative anemia occurs because of hemorrhage or hemolysis. Some find it useful to remember these as the “2 Hs” or alternatively, the “2 Ls” (loss or lysis). In the case of hemorrhage, erythrocytes and the other components of blood escape from the vasculature. Hemorrhage may be acute or chronic or internal or external. Causes of hemorrhage include trauma, abnormal hemostasis, certain forms of parasitism, ulceration, and neoplasia. Thus regenerative anemias usually occur not because of a problem with erythropoiesis but because of a process affecting erythrocytes that have already been released into the blood circulation. However, it is important to note that chronic hemorrhage can deplete the body’s iron stores, leading to iron-deficiency anemia, which may be either regenerative or nonregenerative. A regenerative response may occur when the deficiency is resolving or temporarily compensated (e.g., when hemorrhage ceases, or when a patient suddenly gains access to increased dietary or parenteral iron). Nonregenerative anemias and iron-deficiency anemia specifically are discussed in more detail later in this chapter.
Hemolysis may be intravascular, in which case erythrocytes release their contents, mostly hemoglobin, directly into the blood, or extravascular, in which case macrophages phagocytose erythrocytes, and little or no hemoglobin is released into the blood. Both forms (mostly extravascular hemolysis) occur as part of normal homeostasis and involve pathways to conserve iron and other reusable components in hematopoiesis. However, some diseases are associated with increased destruction of erythrocytes by one, or both, of these mechanisms. The next sections discuss basic mechanisms of intravascular and extravascular hemolysis in health and disease in more detail.
Normal turnover of erythrocytes occurs mainly by extravascular hemolysis, in which senescent erythrocytes are phagocytosed by macrophages in the spleen and to a lesser extent in other organs such as liver (Kupffer cells) and bone marrow. The exact controls are not clear, but factors that likely play a role include the following:
• Exposure of membrane components normally sequestered on the inner leaflet of the cell membrane, particularly phosphatidylserine (this mechanism is also important in apoptosis of other cell types).
Macrophages degrade erythrocytes into reusable constituents, such as iron and amino acids, and the waste product bilirubin. Intravascular hemolysis normally occurs at only extremely low levels. Hemoglobin is a tetramer that, when released from the erythrocyte into the blood, splits into dimers that bind to a plasma protein called haptoglobin. The hemoglobin-haptoglobin complex is taken up by hepatocytes and macrophages. This is the major pathway for handling free hemoglobin. However, free hemoglobin may also oxidize to form methemoglobin, which dissociates to form metheme and globin. Metheme binds to a plasma protein called hemopexin, which is taken up by hepatocytes and macrophages in a similar manner to hemoglobin-haptoglobin complexes. Free heme in the reduced form binds to albumin, from which it is taken up in the liver and converted into bilirubin.
In hemolytic anemia, erythrocytes are destroyed at an increased rate. Whether the mechanism is intravascular or extravascular, or a combination, depends on the specific disease process (specific diseases are discussed later in this chapter). A classic sequela of hemolytic anemias in general is hyperbilirubinemia, which is an increase in the plasma concentration of bilirubin. Bilirubin is a yellow pigment, which explains why hyperbilirubinemia, if severe enough, causes icterus—the grossly visible yellowing of serum or tissue (Fig. 13-9). Icterus of mucous membranes, skin, and other tissue is known as jaundice. Icterus is usually detectable when the plasma bilirubin concentration exceeds 2 mg/dL. Hemolysis is a cause of prehepatic hyperbilirubinemia, but it is important to note that hyperbilirubinemia can also occur as the result of hepatic or posthepatic conditions causing impaired bile flow (cholestasis).
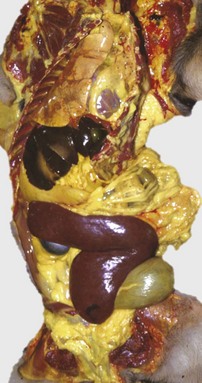
Fig. 13-9 Icterus, immune-mediated hemolytic anemia (IMHA), subcutaneous fat, splenomegaly, spleen, dog.
The marked yellow discoloration of tissues, most strikingly visible in the subcutaneous fat, is from high concentrations of serum bilirubin produced as a result of the hemolytic anemia. (Courtesy Dr. J.A. Ramos-Vara, College of Veterinary Medicine, Michigan State University; and Noah’s Arkive, College of Veterinary Medicine, The University of Georgia.)
Laboratory findings and clinical observations may point to a specific mechanism of hemolysis. In patients with extravascular hemolytic anemia, increased destruction of erythrocytes by splenic macrophages often results in splenomegaly (Fig. 13-10). Splenomegaly may also occur because of other conditions, as discussed elsewhere in this chapter. Intravascular hemolysis is grossly evident as hemoglobinemia (pink-tinged plasma or serum), if the concentration of extracellular hemoglobin is greater than approximately 50 mg/dL. Haptoglobin is saturated with dimeric hemoglobin at a concentration of approximately 150 mg/dL. When haptoglobin is saturated, any remaining free hemoglobin has a low enough molecular weight to pass through the renal glomerular filter into the urine (hemoglobinuria), imparting a pink or red discoloration to the urine. Thus extracellular hemoglobin can cause gross discoloration of the plasma, where it is bound to haptoglobin, before becoming grossly visible in urine. The half-life of haptoglobin is markedly decreased when bound to hemoglobin, so when large amounts of haptoglobin-hemoglobin complex are formed, the concentration of haptoglobin in the blood decreases and hemoglobin can pass through the glomerulus at even lower concentrations. Hemoglobinuria is a contributing factor in the renal tubular necrosis (hemoglobinuric nephrosis) that often occurs in cases of acute intravascular hemolysis (see Chapter 11). A similar lesion occurs in the kidneys of individuals with marked muscle damage and resulting myoglobinuria.
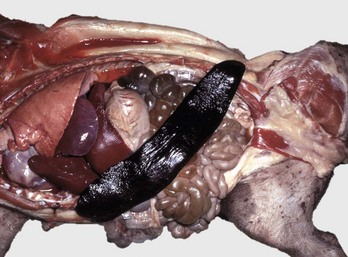
Fig. 13-10 Splenomegaly, fatal hemolytic anemia, Mycoplasma suis, pig.
The spleen is extremely enlarged, meaty, and congested. (Courtesy College of Veterinary Medicine, University of Illinois.)
Hemoglobinuria cannot be distinguished grossly from hematuria (RBCs in the urine) or myoglobinuria, and both hemoglobin and myoglobin cause a positive reaction for “blood protein” on urine test strips. Comparing the colors of the plasma and the urine may be informative. In contrast to hemoglobin, myoglobin causes gross discoloration of the urine before the plasma is discolored. This is because myoglobin is a low-molecular-weight monomer, freely filtered by the glomerulus, and does not bind plasma proteins to a significant degree. Hematuria can be distinguished from hemoglobinuria on the basis of microscopic examination of urine sediment (RBCs are present in cases of hematuria).
CBC data are often useful in illuminating hemolytic mechanisms. Total hemoglobin concentration is conventionally measured by lysing all the erythrocytes and measuring the hemoglobin in solution spectrophotometrically. The value for mean cell hemoglobin (MCH) is calculated based on the total hemoglobin concentration and the concentration of erythrocytes. The value for MCHC is calculated based on the MCH and MCV. Thus although erythrocytes do not actually contain supranormal concentrations of hemoglobin, an excess of extracellular hemoglobin may cause an artifactual increase in the calculated MCH and MCHC. It is important to remember that similar increases may also occur artifactually because of positive interference with spectrophotometric measurement of hemoglobin such as occurs with lipemia.
Extravascular hemolysis also often produces characteristic CBC abnormalities that may reflect the mechanism of disease. For example, spherocytosis and autoagglutination are hallmarks of IMHA. Spherocytes form when macrophages (mainly in the spleen) phagocytose part of an erythrocyte plasma membrane bound with autoantibody (Fig. 13-11). The remaining portion of the RBC assumes a spherical shape, thus preserving maximal volume. This change in shape results in decreased deformability of the cells. Erythrocytes must be extremely pliable to traverse the splenic red pulp and sinusoidal walls (Fig. 13-12); spherocytes therefore tend to be retained in the spleen in close association with macrophages with risk of further injury and eventual destruction. In the dog, spherocytes appear smaller than normal and have uniform staining (Fig. 13-13, A), in contrast to normal RBCs, which have a region of central pallor imparted by their biconcave shape. This difference in staining between spherocytes and normal RBCs is not consistently discernible in many other domestic animals (including horses, cattle, and cats), whose erythrocytes differ from those of the dog in that they are smaller and have less pronounced biconcavity and therefore less pronounced central pallor. Autoagglutination occurs because of cross-linking of antibodies bound to erythrocytes. Autoagglutination is evident microscopically as clusters of erythrocytes, and macroscopically as blood with a grainy consistency (Fig. 13-13, B). Autoagglutination may also result in a falsely increased MCV and decreased RBC concentration when clustered cells are mistakenly counted as single cells by automated hematology analyzers.
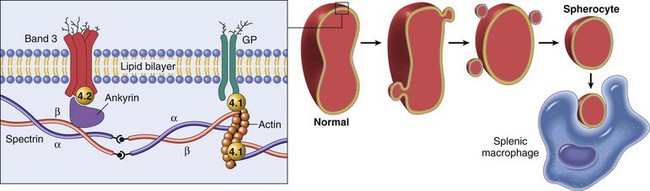
Fig. 13-11 Schematic representation of the mechanisms of spherocytosis and extravascular hemolysis (EVH).
Hereditary spherocytosis occurs in humans (as shown here) because of mutations that weaken the connections between cytoskeletal and membrane proteins. In cases of immune-mediated hemolytic anemia (IMHA), the most frequent cause of spherocytes in animals, the underlying disease mechanism is different—spherocytes form when portions of the erythrocyte membrane bound with autoantibody are phagocytosed by macrophages—but the net result (spherocytosis and EVH) is similar to what is shown here. GP, Glycoprotein. (From Kumar V, Abbas AK, Fausto N, et al: Robbins & Cotran pathologic basis of disease, ed 8, Philadelphia, 2009, Saunders.)
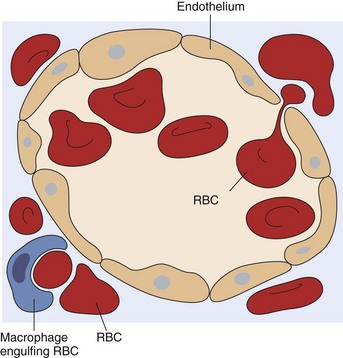
Fig. 13-12 Schematic representation of a splenic sinusoid.
A red blood cell (RBC) is in the process of squeezing from the red pulp cords into the sinus lumen. Note the degree of deformability required for RBCs to pass through the wall of the sinus. (From Kumar V, Abbas AK, Fausto N: Robbins & Cotran pathologic basis of disease, ed 7, Philadelphia, 2005, Saunders.)
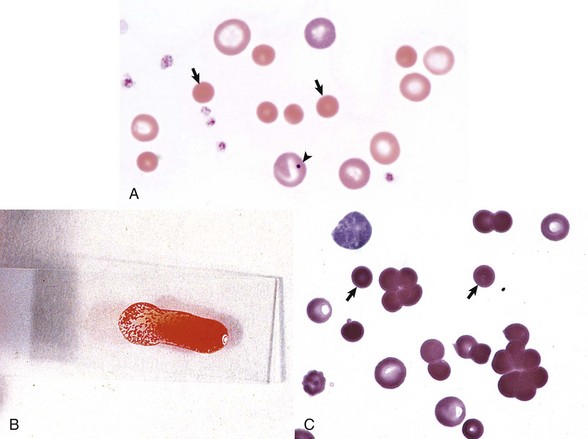
Fig. 13-13 Immune-mediated hemolytic anemia (IMHA), canine blood, dog.
A, Spherocytosis. Numerous spherocytes (arrows) and several polychromatophilic cells, one of which (arrowhead) contains a nuclear remnant known as a Howell-Jolly body, are visible. Wright’s stain. B, Autoagglutination. Note the grossly visible agglutination. C, Spherocytosis and agglutination. Note the spherocytes (arrows) and microscopic agglutination of erythrocytes in a dog that received a blood transfusion. These abnormalities presumably indicate immune-mediated destruction of the donor erythrocytes. Wright’s stain. (A courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee. B and C from Harvey JW: Atlas of veterinary hematology: blood and bone marrow of domestic animals, Philadelphia, 2001, Saunders.)
Oxidative damage to erythrocytes occurs when normal antioxidative pathways that generate reducing agents (such as NADH, NADPH, and reduced glutathione) are compromised or overwhelmed and can result in hemolytic anemia, abnormal hemoglobin function, or both. Hemolysis caused by oxidative damage may be extravascular or intravascular, or a combination (predominant forms of hemolysis are discussed further in the section on specific diseases). Evidence of oxidative damage to erythrocytes may be apparent on examination of a blood smear, or even on gross physical examination. Heinz bodies are foci of denatured globin that interact with the erythrocyte membrane. They are usually subtly evident on routine Wright’s-stained blood smears as pale circular inclusions or blunt, rounded protrusions of the cell margin and are readily discernible on smears stained with new methylene blue (Fig. 13-14). Cats are particularly susceptible to Heinz body formation and may have low numbers of small Heinz bodies normally. There is no unanimity of opinion, but some clinical pathologists believe that the presence of Heinz bodies in up to 10% of all RBCs in cats is within normal limits. This predisposition is believed to reflect unique features of the feline erythrocyte, whose hemoglobin has more sulfhydryl groups (preferential sites for oxidative damage) than do erythrocytes of other species and may also have lower intrinsic reducing capacity. It is also possible that the feline spleen, which is nonsinusoidal, does not have as efficient a “pitting” function as does a sinusoidal spleen, such as that of the dog (splenic structure and function are discussed in more detail later in this chapter). Eccentrocytes, evident as erythrocytes in which one side of the cell has increased pallor, are another manifestation of oxidative damage. They form because of cross-linking of membrane proteins, with adhesion of opposing areas of the cell’s inner membrane leaflet, and displacement of most of the hemoglobin toward the other side (Fig. 13-15). Oxidative insult may also result in conversion of hemoglobin (iron in the Fe2+ state) to methemoglobin (iron in the Fe3+ state), which is incapable of binding oxygen. Methemoglobin is produced normally in small amounts but reduced back to oxyhemoglobin by the enzyme cytochrome-b5 reductase (also known as methemoglobin reductase). Methemoglobinemia results when methemoglobin is produced in excessive amounts (because of oxidative insult) or when the normal pathways for maintaining hemoglobin in the Fe2+ state are impaired (as in cytochrome-b5 reductase deficiency). When present in sufficiently high concentration (approximately 10% of total hemoglobin), methemoglobin imparts a grossly discernible chocolate color to the blood.
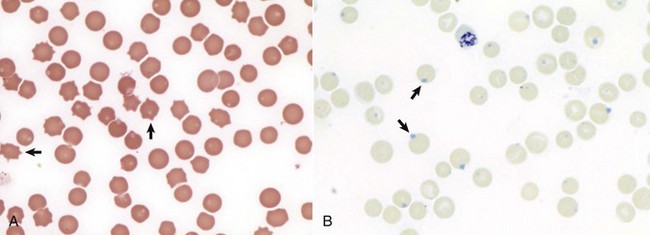
Fig. 13-14 Heinz bodies, blood smears.
A, Feline blood smear. With routine staining, Heinz bodies appear as pale circular intraerythrocytic inclusions that may protrude (arrows) from the margin of the cell. Wright’s stain. B, Canine blood smear. Using a supravital stain, Heinz bodies are blue inclusions (arrows) and easier to see. New methylene blue stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
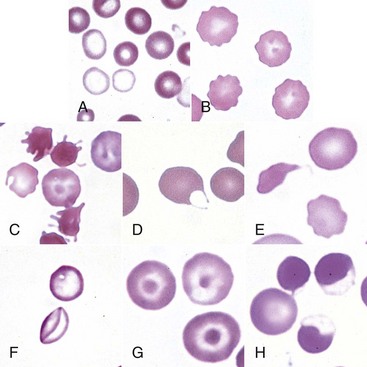
Fig. 13-15 Common erythrocyte morphologic abnormalities.
A, Blood from a dog with a microcytic hypochromic iron-deficiency anemia was mixed with an equal volume of blood from a normal dog before blood smear was prepared. Because the hypochromic (pale-staining) cells are leptocytes shown in F, they have diameters similar to normal cells even though they are microcytic cells. Wright-Giemsa stain. B, Echinocytes appear as erythrocytes with scalloped borders; consequently, the old term “crenation” from Latin meaning “notched” is used. Wright-Giemsa stain. C, Three acanthocytes with irregularly spaced, variably sized spicules in blood from a dog with hemangiosarcoma. Wright-Giemsa stain. D, A keratocyte, exhibiting what appears to be a ruptured “vesicle” in blood from a cat with hepatic lipidosis. Wright-Giemsa stain. E, A schistocyte (left), discocyte (top), and echinocyte (bottom) in blood from a dog with disseminated intravascular hemolysis. Wright-Giemsa stain. F, Two thin flat hypochromic-appearing erythrocytes (leptocytes), with increased membrane-to-volume ratios, are present in blood from a dog with severe iron-deficiency anemia. The bottom leptocyte is folded. Wright-Giemsa stain. G, Three codocytes in blood from a Cairn terrier with a regenerative anemia and hepatic hemochromatosis secondary to pyruvate kinase deficiency. These erythrocytes exhibit a central density or “bull’s-eye” and are often referred to as target cells. Wright-Giemsa stain. H, Three eccentrocytes and a discocyte (left) in blood from a dog with oxidant injury induced by the administration of acetaminophen. The cell at top center appears spherical with a small tag of cytoplasm and may be referred to as a pyknocyte. Wright-Giemsa stain. (From Harvey JW: Atlas of veterinary hematology: blood and bone marrow of domestic animals, Philadelphia, 2001, Saunders.)
Nonregenerative anemias are characterized by a lack of reticulocytosis on the CBC; however, reticulocytosis does not occur in horses even in the context of regeneration. Most often this is a result of decreased production in the marrow (i.e., erythroid hypoplasia). Erythrocytes circulate for a long time, so anemias caused by decreased production tend to develop slowly. The most common form of nonregenerative anemia is known as anemia of inflammation or anemia of chronic disease. In this form of anemia, RBCs are decreased in number but are typically normal in size and hemoglobin concentration (so-called normocytic, normochromic, anemia). It has long been known that patients with inflammatory or other chronic disease often become anemic and that this condition is associated with increased iron stores in the bone marrow. Sequestration of iron may be a bacteriostatic evolutionary adaptation because many bacteria require iron as a cofactor for growth. In recent years, investigators have begun to elucidate the molecular mechanisms underlying anemia of inflammation. Hepcidin, an acute phase protein synthesized in the liver that was first identified as an antimicrobial peptide, is a key mediator that acts by limiting iron availability. Hepcidin expression increases with inflammation, infection, or iron overload and decreases with anemia or hypoxia. Hepcidin exerts its effects by causing functional iron deficiency. It binds to and causes the degradation of the cell surface iron efflux molecule, ferroportin, thus inhibiting both absorption of dietary iron from the intestinal epithelium and export of iron from macrophages and hepatocytes into the plasma (Fig. 13-16).
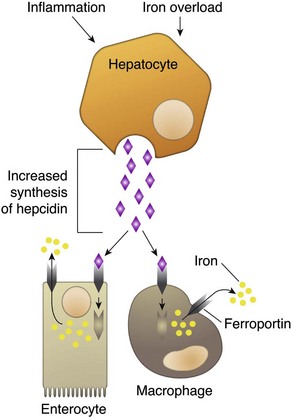
Fig. 13-16 Mechanism of action of hepcidin.
Binding of hepcidin to ferroportin causes its internalization and degradation, thus inhibiting efflux of iron into the plasma. (Courtesy Dr. J.F. Zachary, College of Veterinary Medicine, University of Illinois.)
Anemia of inflammation involves factors besides decreased iron availability. Inflammatory cytokines are likely to inhibit erythropoiesis by oxidative damage to and triggering apoptosis of developing erythroid cells, by decreasing expression of Epo and stem-cell factor, and by decreasing expression of Epo receptors. In addition, experimentally induced sterile inflammation in cats resulted in shortened erythrocyte survival, indicating that anemia of inflammation is likely also a function of increased erythrocyte destruction.
True iron deficiency has long been recognized as a cause of anemia. In domestic animals, iron deficiency occurs most commonly because of chronic blood loss, and thus loss of iron in the form of hemoglobin, and less frequently because of nutritional deficiency. Although iron deficiency often results in nonregenerative anemia, it is not always so, especially when nutritional iron is not a limiting factor. Many underlying conditions can cause hemorrhage-induced iron deficiency anemia, including primary or secondary gastrointestinal (GI) disease (e.g., hookworms, neoplasia, ulceration) and marked ectoparasitism. The classic hematologic picture with iron deficiency is microcytic, hypochromic anemia. Microcytosis and hypochromasia are indicated on the CBC by abnormally low MCV and MCHC values, respectively. Microcytosis and hypochromasia may also be discernible on review of a blood smear as RBCs that are abnormally small or paler-staining because of their subnormal hemoglobin concentration, respectively. However, microscopic examination is not a reliable means of detection, especially in the case of mild abnormalities. Low hemoglobin concentration is believed to contribute to microcytosis because one of the feedback mechanisms signaling erythroid cells to stop dividing is reaching a threshold hemoglobin concentration. Abnormally low hemoglobin concentration during erythropoiesis thus results in extra cell divisions and smaller erythrocytes. Microcytosis usually develops before hypochromasia.
Other causes of decreased erythropoiesis include the following:
• Decreased hormonal stimulation (e.g., chronic renal failure)
• Infection of erythropoietic cells
• Toxic insult to the bone marrow
• Other disease involving the bone marrow
Specific examples of diseases causing nonregenerative anemia by these mechanisms are discussed later in this chapter. It is important to point out that nonregenerative anemia is not always caused by decreased erythropoiesis. For example, IMHA is typically strongly regenerative, but there are also nonregenerative forms of IMHA. Bone marrow findings in dogs with severe nonregenerative IMHA range from a complete absence of erythropoiesis, known as pure red cell aplasia, to erythroid hyperplasia in a majority of patients in one study. The latter situation is an example of ineffective hematopoiesis (in this case, ineffective erythropoiesis), in which cells are produced at abnormal or increased rate but are destroyed, presumably because of an immune-mediated mechanism, before they enter the circulation.
Neutropenia: Neutropenia refers to a decrease in the concentration of neutrophils in circulating blood. Neutropenia may be caused by decreased production, increased destruction, altered distribution, demand for neutrophils in inflamed tissue that exceeds the rate of granulopoiesis, or inherited disease. Decreased production is evident on bone marrow examination as granulocytic hypoplasia. This usually results from an insult that affects multiple hematopoietic lineages, such as chemical insult, radiation, neoplasia, infection, and fibrosis, but may also be caused by a process that preferentially targets granulopoiesis. Immune-mediated neutropenia is a rare but recognized condition in domestic animals. Bone marrow findings range from granulocytic aplasia or hypoplasia to hyperplasia, depending on where the cells under immune attack are in their differentiation programs. Neutropenia with no evidence of decreased production and in which other causes of neutropenia have been excluded may be a result of destruction of neutrophils before they leave the bone marrow, a condition known as ineffective granulopoiesis. Like other forms of ineffective hematopoiesis, this condition is often presumed to be immune mediated; in cats this condition may occur as a result of infection of hematopoietic cells with the feline leukemia virus (FeLV).
In marked contrast to erythrocytes, neutrophils have a very short lifespan in circulation. Once released from the bone marrow, a neutrophil is only in the bloodstream for hours before migrating into the tissues. The fate of neutrophils after they leave the bloodstream in normal conditions (i.e., not in the context of inflammation) is poorly understood. They migrate into the GI and respiratory tracts, liver, and spleen and may be lost through mucosal surfaces or undergo apoptosis and be phagocytosed by macrophages. When neutrophil production ceases, a reserve of mature neutrophils in the bone marrow (discussed in more detail later) may be adequate to maintain normal numbers of circulating neutrophils for a few days; however, after the bone marrow storage pool is depleted, neutropenia rapidly ensues.
Neutrophils within the blood vasculature are in two compartments: a circulating pool, consisting of those cells flowing freely in the blood, and a marginated pool, consisting of those cells transiently associated with the endothelial surface. (In reality, neutrophils are constantly shifting between these two pools, but the proportion of cells in either pool normally remains fairly constant in any given species.) Circulating neutrophils are part of the blood sample collected during routine venipuncture and are thus counted in the CBC, whereas marginated neutrophils are not. Pseudoneutropenia refers to the situation in which there are an increased proportion of neutrophils in the marginated pool. This may occur because of decreased blood flow or in response to stimuli, such as endotoxemia, that increase expression of molecules promoting interaction between neutrophils and endothelial cells.
Neutropenia may also result from increased demand for neutrophils in the tissue. How rapidly such a situation develops depends not only on the magnitude of the inflammatory stimulus but also on the reserve of postmitotic neutrophils in the bone marrow. The size of this reserve, or storage pool, is species dependent. In dogs, this pool contains the equivalent of 5 days normal production of neutrophils. Cattle represent the other extreme in that they have a small storage pool and thus are predisposed to becoming neutropenic more easily than are dogs. Horses and cats are somewhere between the two extremes, closer to cattle and dogs, respectively. It stands to reason that the clinical significance of neutropenia because of a supply and demand imbalance is also species dependent. In dogs, neutropenia as a result of inflammation is an alarming finding because it is evidence of a massive tissue demand for neutrophils that has exhausted the patient’s storage pool and is exceeding the rate of granulopoiesis in the bone marrow, whereas in cows, neutropenia is commonly noted in a wide range of conditions involving acute inflammation and does not necessarily indicate an overwhelming demand.
Eosinopenia/Basopenia: In many laboratories, CBC reference values for eosinophils and basophils are as low as zero cells per microliter, precluding detection of eosinopenia or basopenia. When detectable, eosinopenia is often part of a stress (glucocorticoid-mediated) leukogram.
Thrombocytopenia: Thrombocytopenia refers to a decrease in the concentration of circulating platelets. Mechanisms of thrombocytopenia include decreased production, increased destruction, increased consumption, altered distribution, and hemorrhage. Decreased production may occur because of a condition affecting cells of multiple hematopoietic lineages, including platelets, or to one specifically depressing thrombopoiesis. In either case, decreased thrombopoiesis is evident on examination of a bone marrow sample as megakaryocytic hypoplasia. General causes of decreased hematopoiesis outlined earlier in the sections on anemia and neutropenia also apply to thrombocytopenia. Specific diseases causing thrombocytopenia are covered later in this chapter. Immune-mediated thrombocytopenia is a fairly common disease in dogs and may also occur in other species. Increased consumption of platelets is a hallmark of disseminated intravascular coagulation (DIC), a syndrome in which hypercoagulability leads to increased consumption of both platelets and coagulation factors in the plasma, with subsequent hypocoagulability and susceptibility to bleeding. The spleen normally contains a significant proportion of total platelet mass (up to one-third in some species), and abnormalities involving the spleen may result in changes in the number of circulating platelets. For example, splenic congestion may result in thrombocytopenia, and splenic contraction may result in thrombocytosis. Acute hemorrhage may result in mild to moderate thrombocytopenia. Potential mechanisms of thrombocytopenia caused by hemorrhage include loss and consumption. Of course, thrombocytopenia can also be a cause of hemorrhage. In the absence of other complicating factors, marked to severe thrombocytopenia (less than 50,000 platelets/µL) is more likely to be the cause of rather than the result of bleeding. Megakaryocytic hyperplasia on examination of a bone marrow sample is evidence of a regenerative thrombopoietic response, and an increase in the mean platelet volume (MPV) value on the CBC often accompanies such a response.
Lymphopenia: Lymphopenia refers to a decrease in the concentration of lymphocytes in blood circulation. It is a common CBC finding in sick animals. Usually the precise mechanism of lymphopenia is not clear. It is often presumed to be mediated at least in part by endogenous glucocorticoid excess. Lymphopenia may occur because of various mechanisms, including altered distribution of lymphocytes (increased trafficking of lymphocytes to, and decreased egress from, lymphoid tissues), lymphotoxicity (direct damage to lymphocytes or suppression of lymphopoiesis) of therapeutic or infectious agents, loss of lymphocyte-rich lymphatic fluid, or congenital disorders. Normal lymphocyte trafficking may be altered because of disruption of the normal architecture of lymphoid tissue (e.g., because of neoplasia or inflammation), or in response to cytokine signals. Glucocorticoid excess may cause lymphopenia via redistribution from the blood to lymphoid tissue, or via direct lymphotoxic effects. Anticancer treatments (chemotherapy or radiotherapy) and immunosuppressive drugs may also be lymphotoxic. Some hereditary immunodeficiencies (such as severe combined immunodeficiency or thymic aplasia) can cause lymphopenia.
Erythrocytosis: An increase in the measured red cell mass above the normal range (i.e., the opposite of anemia) is known as erythrocytosis. The term polycythemia is often used interchangeably with erythrocytosis, but technically and for the purposes of this chapter, polycythemia refers to a specific type of leukemia called primary erythrocytosis or polycythemia vera. Relative erythrocytosis occurs most frequently because of dehydration, when the decreased proportion of water in the blood results in hemoconcentration. A similar situation may occur as a result of epinephrine-mediated splenic contraction. Erythrocytosis caused by splenic contraction occurs to the most pronounced degree in the horse and may occur to a lesser extent in other species. In both of these cases, the total RBC mass is not in fact increased but appears to be so because of other factors. Secondary erythrocytosis refers to a true, Epo-mediated increase in red cell mass, either as an appropriate response to hypoxemia (such as may be seen in patients with cardiopulmonary disease and severely impaired oxygenation, or cardiovascular defects causing right-to-left shunting) or in rare cases an Epo-secreting tumor (inappropriate secondary erythrocytosis). Polycythemia vera is diagnosed based on a marked increase in red cell mass (hematocrit in normally hydrated dogs ranges from 65% to >80%), absence of hypoxemia, absence of other tumors, and normal or decreased concentration of plasma erythropoietin concentration. Absolute erythrocytosis, whether primary or secondary, causes increased viscosity of the blood and resulting impairment of blood flow and distention of the microvasculature. Affected individuals are at increased risk of tissue hypoxia and thrombosis or hemorrhage. Related clinical signs (hyperviscosity syndrome) may include erythematous mucous membranes (Fig. 13-17) and prolonged capillary refill time, prominent scleral vessels, evidence of thrombosis or hemorrhage, and secondary signs (e.g., neurologic and cardiovascular) related to specific organ systems affected.
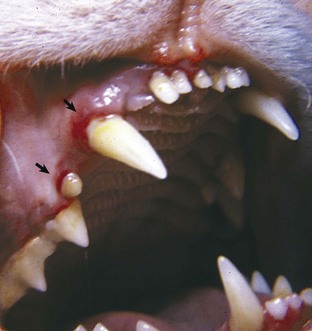
Fig. 13-17 Absolute erythrocytosis, hyperviscosity syndrome, erythematous mucous membranes, cat.
Erythema of mucous membranes is one of the signs associated with hyperviscosity syndrome. In this case, the oral mucous membranes are deeper red (arrows) than normal because of an abnormally high concentration of erythrocytes and associated sludging of blood. Hyperviscosity syndrome may also occur as the result of increased plasma immunoglobulin concentration. (Courtesy Dr. C. Patrick Ryan, Veterinary Public Health, Los Angeles Department of Health Services; and Noah’s Arkive, College of Veterinary Medicine, University of Georgia.)
Neutrophilia: Neutrophilia occurs in response to a number of different stimuli, which are not mutually exclusive. Major mechanisms of neutrophilia are shown in Fig. 13-18. Understanding the CBC findings characteristic of these responses is an important part of clinical veterinary medicine. Inflammation can result in neutropenia, as discussed earlier, or neutrophilia, as discussed next. However, before moving on to a discussion of inflammatory neutrophilia and the so-called left shift, it is important to mention two other common causes of neutrophilia.
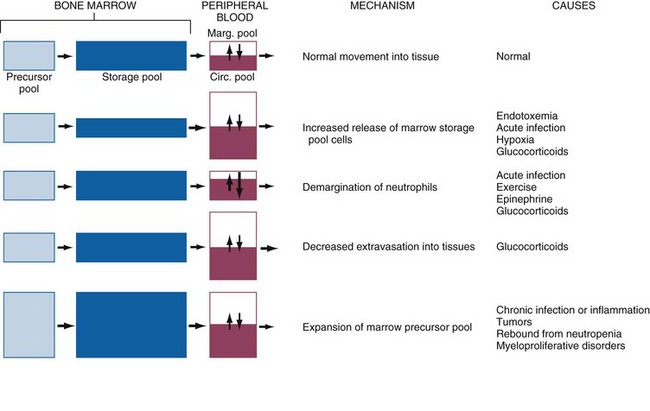
Fig. 13-18 Schematic illustration of the mechanisms of neutrophilic leukocytosis.
Neutrophils and their precursors are distributed in five pools: a bone marrow precursor pool, which includes progenitor cells and more committed, mitotically active precursors; a bone marrow storage pool, consisting of mitotically inactive mature and slightly immature neutrophils (band forms); a peripheral blood marginating pool (Marg. pool); a peripheral blood circulating pool (Circ. pool); and a tissue pool. The relative size of each pool is represented by the size of its corresponding box. The peripheral blood neutrophil count measures only the circulating peripheral blood pool, which can be enlarged by increased release of cells from the marrow storage pool, increased demargination, diminished extravasation into tissue, or expansion of the marrow precursor cell pool. (Modified from Finch SC: Hematology, ed 3, New York, 1983, McGraw-Hill.)
Glucocorticoid excess, either because of endogenous production or exogenous administration, results in a CBC pattern known as the stress leukogram, characterized by mature neutrophilia (i.e., increased concentration of segmented neutrophils), lymphopenia, and, especially in dogs, monocytosis. Eosinopenia is another feature of the stress leukogram, although in many situations this is inapparent because the normal reference values for eosinophils are so low (in some laboratories, the lower reference value is zero). Mechanisms contributing to glucocorticoid-mediated neutrophilia include the following:
• Increased release of mature neutrophils from the bone marrow storage pool.
• Decreased margination of neutrophils within the vasculature, with a resulting increase in the circulating pool.
• Decreased migration of neutrophils from the bloodstream into tissues.
The magnitude of neutrophilia tends to be species dependent, with dogs having the most pronounced response (up to 35,000 cells/µL) and in decreasing order of responsiveness, cats (30,000 cells/µL), horses (20,000 cells/µL), and cows (15,000 cells/µL) less marked responses. With long-term glucocorticoid excess, neutrophil numbers tend to normalize, whereas lymphopenia persists.
Epinephrine release results in a different pattern, known as physiologic leukocytosis, characterized by mature neutrophilia (like the glucocorticoid response) and lymphocytosis (unlike the glucocorticoid response). This phenomenon is short lived (<1 hour). Neutrophilia occurs primarily because of a shift of cells from the marginated to the circulating pool. Physiologic leukocytosis is common in cats (especially when they are highly stressed during blood collection) and horses, less common in cattle, and uncommon in dogs.
Of course, neutrophilia may also indicate inflammation, and inflammatory stimuli of varying magnitude and duration produce different patterns of neutrophilia. A classic hematologic finding in patients with increased demand for neutrophils is the presence of immature forms in the blood, known as a “left shift.” Not all inflammatory responses have a left shift, but the presence of a left shift almost always signifies active demand for neutrophils in the tissue. The magnitude of a left shift is assessed by the number of immature cells and their degree of immaturity. The mildest form is characterized by increased numbers of band neutrophils, the immediate predecessor to the segmented neutrophil normally found in circulation, with increasingly severe forms involving progressively immature predecessors. A left shift is considered orderly if the number of immature neutrophils in circulation decreases as they become progressively immature. The term degenerative left shift is sometimes used to describe cases where the number of immature forms exceeds the number of segmented neutrophils. As with glucocorticoid-mediated neutrophilia, the typical magnitude of neutrophilia caused by inflammation varies by species, with dogs having the most pronounced response.
Marked neutrophilia may also occur because of an inherited condition known as leukocyte adhesion deficiency (LAD), in which leukocytes lack normal expression of an adhesion molecule required for migration from the bloodstream into the tissue. This condition is described further in the section on specific diseases.
It might be useful to think of neutrophil kinetics in terms of a producer-consumer model in which the bone marrow is the factory, and the tissues (where the neutrophils eventually go) are the customers. The bone marrow storage pool is the factory inventory, and the neutrophils in the bloodstream are in delivery to the customer. Within the blood vessels, circulating neutrophils are on the highway, and marginated neutrophils are temporarily pulled off to the side of the road. During health, there is an even flow of neutrophils from the factory to the customer. Thus the system is in steady state, and neutrophil numbers remain relatively constant and within the normal range. However, disease states may perturb this system at multiple levels. Decreased granulopoiesis is analogous to a factory working below normal production level. Ineffective granulopoiesis is analogous to goods produced at a normal to increased rate but that are damaged during manufacturing and never leave the factory. A left shift is analogous to the factory meeting increased customer demand by shipping out unfinished goods. Cases of persistent, established inflammation are characterized by bone marrow granulocytic hyperplasia and mature neutrophilia, analogous to a factory that has had time to adjust to increased demand and is meeting it more efficiently by increasing its output.
Eosinophilia/Basophilia: Eosinophilia and, less commonly, basophilia (which, when present, often occurs concurrently with eosinophilia) may occur in response to parasitism, as part of an allergic response, as a paraneoplastic abnormality (because of production of cytokines by neoplastic cells or other inflammatory cells reacting to the neoplasm), as an atypical response to nonparasitic infectious disease, or in rare idiopathic conditions (e.g., hypereosinophilic syndrome).
Thrombocytosis: Thrombocytosis, or concentration of platelets in the blood above normal reference values, is a relatively common nonspecific finding in veterinary patients. In the vast majority of cases, thrombocytosis is reactive—a response to another, often apparently unrelated, disease process. Examples of conditions associated with reactive thrombocytosis include inflammatory and infectious diseases, iron deficiency, hemorrhage, endocrinopathies, and neoplasia. Factors that may contribute to reactive thrombocytosis include increased plasma concentration of thrombopoietin, inflammatory cytokines (e.g., IL-6), or catecholamines. Thrombocytosis may also occur as part of a regenerative response in patients recovering from thrombocytopenia, or as a result of redistribution because of splenic contraction (especially in horses) or splenectomy. In these cases, thrombocytosis is transient. In the case of splenectomy, thrombocytosis may be marked, but normalizes after several weeks. Because the body’s total platelet mass regulates thrombopoiesis and a significant portion of the platelet mass is normally in the spleen, it makes sense that splenectomized animals develop thrombocytosis. However, the reason that the number of circulating platelets normalize in these individuals in the weeks after splenectomy is not clear. There is also a rare form of leukemia known as essential thrombocythemia, which is characterized by marked thrombocytosis.
Lymphocytosis: Lymphocytosis refers to an increase in the concentration of lymphocytes in blood circulation. There are numerous causes of lymphocytosis. Young animals normally have higher concentrations of lymphocytes than older animals, and normal healthy young animals may have counts that exceed adult reference values. As discussed earlier in the section on neutrophilia, lymphocytosis is also a feature of epinephrine-mediated physiologic leukocytosis, resulting from redistribution of lymphocytes into the blood circulating pool. Epinephrine-mediated lymphocytosis may be more marked than neutrophilia, particularly in cats (lymphocyte counts of >20,000/µL are not uncommon). Antigenic stimulation may result in lymphocytosis, even in rare cases in marked lymphocytosis (up to approximately 30,000 cells/µL in dogs, and 40,000/µL in cats); however, this is not usually the case, even when there is clear evidence of increased immunologic activity in lymphoid tissues. In cases of antigenic stimulation, it is common for a minority of lymphocytes to have a “reactive” morphology—larger in size than small, mature lymphocytes, and with more abundant, deeply basophilic cytoplasm and incompletely condensed chromatin (Fig. 13-19). Just as glucocorticoid excess can cause lymphopenia, glucocorticoid deficiency (hypoadrenocorticism) can cause lymphocytosis, or lack of lymphopenia under conditions of stress that typically result in glucocorticoid-mediated lymphopenia.
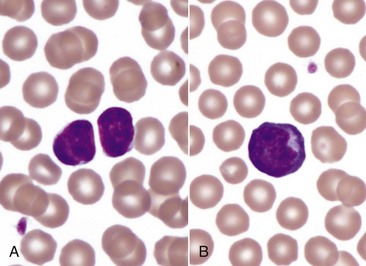
Fig. 13-19 Lymphocytosis (B), lymphocytes, canine blood smear.
A, Small lymphocytes, the predominant type of lymphocyte in the blood under normal conditions. B, A reactive lymphocyte, characterized by mildly increased size and an increased amount of basophilic cytoplasm, from a recently vaccinated 16-week-old dog. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
A condition known as persistent lymphocytosis (PL) occurs in approximately 30% of cattle infected with the bovine leukemia virus (BLV). The condition is defined as an increase in the blood concentration of lymphocytes above the reference interval for at least 3 months. This form of lymphocytosis is a nonneoplastic proliferation (hyperplasia) of B lymphocytes. In the absence of other disease, cattle with PL are asymptomatic. However, cattle infected with BLV, especially those animals with PL, are at increased risk to develop B lymphocyte lymphoma. (BLV and lymphoma in cattle are discussed in more detail later in this chapter in the section on Hematopoietic Neoplasia.) As in most other forms of benign lymphocytosis, the morphology of most lymphocytes is within normal limits in cattle with PL.
Secondary Abnormal Structure or Function of Blood Cells: The preceding section focused on abnormalities in the number of blood cells. There are also various acquired and congenital conditions involving abnormal structure or function of blood cells. This section briefly discusses abnormal blood cell structure or function occurring secondary to other underlying disease. Primary disorders of blood cells are discussed later in the chapter in the section on specific diseases,
Morphologic abnormalities detected on routine microscopic examination of blood smears may provide important clues about underlying disease processes. Poikilocytosis is a broad term referring to the presence of abnormally shaped erythrocytes in circulation. Web Table 13-1 lists conditions associated with and mechanisms involved in the formation of a number of specific types of erythrocyte morphologic abnormalities, and Fig. 13-15 shows some examples.
WEB TABLE 13-1
Common Erythrocyte Morphologic Abnormalities
| Term | Description | Condition(s) Commonly Associated with Increased Numbers of Abnormal RBCs |
| Macrocyte | Abnormally large | Regenerative anemia FeLV infection (cats) Congenital in some poodle dogs |
| Microcyte | Abnormally small | Iron deficiency Portosystemic shunts Normal in Akita and Shiba dogs |
| Polychromasia | Bluish color (Figs. 13-5, 13-7) | Reticulocytosis (regenerative anemia) |
| Basophilic stippling | Fine punctate inclusions (Fig. 13-7) | Lead toxicity Regenerative anemia (especially ruminants) |
| Howell-Jolly body (nuclear remnant) | Small, round, blue-black inclusion, usually off-center (Fig. 13-13) | Regenerative anemia Splenic dysfunction |
| Hypochromasia | Increased pallor and deformability (Fig. 13-15) | Iron deficiency |
| Heinz body | Pale round inclusion, may bulge from cell margin; stains blue with new methylene blue (Fig. 13-14) | Oxidative damage |
| Poikilocyte | Nonspecific term for shape abnormality—specific types of poikilocytosis listed below | See below |
| Spherocyte | Appears abnormally small with uniform staining (Figs. 13-11 and 13-13) | Extravascular hemolysis |
| Schistocyte | Small, irregular RBC fragment, often crescent shaped (Fig. 13-15) | Microangiopathies (e.g., DIC, hemangiosarcoma, glomerulopathy) Increased RBC deformability (e.g., iron deficiency) |
| Acanthocyte | Few irregular projections (Fig. 13-15) | Abnormal lipid metabolism (e.g., liver disease) Microangiopathies (as per schistocytes) Increased RBC deformability (as per schistocytes) Common normal finding in young goats |
| Echinocyte | Many relatively uniform projections (Fig. 13-15) | Usually in vitro artifact Normal in pigs |
| Eccentrocyte | Eccentric staining (Fig. 13-16) | Oxidative damage |
| Leptocyte | Thin, often hypochromic and/or folded (Fig. 13-15) | Iron deficiency |
| Codocyte (“target cell”) | Area of dense staining within central pallor (Fig. 13-15) | Liver disease |
| Keratocyte | Intact or ruptured “blister” (Fig. 13-15) | Iron deficiency Conditions causing schistocytosis and/or acanthocytosis |
DIC, Disseminated intravascular coagulation; FeLV, feline leukemia virus; RBC, red blood cell.
Similarly the acquired neutrophil morphologic abnormality known as toxic change (Fig. 13-20) reflects accelerated production of neutrophils as part of the inflammatory response. Features of toxic change include increased cytoplasmic basophilia, the presence of small blue-gray cytoplasmic inclusions known as Döhle bodies (often noted incidentally in cats) and in more severe cases, cytoplasmic vacuolation. Although not associated with impaired neutrophil function, toxic change occurs during granulopoiesis and thus is technically a form of dysplasia (e.g., Döhle bodies are foci of aggregated endoplasmic reticulum). Toxic change may accompany any inflammatory response, but in general the more marked the toxic change, the higher the index of suspicion for infection or endotoxemia. Other secondary changes to neutrophils may not be evident morphologically. For example, studies in humans and dogs have shown that individuals with cancer have abnormal neutrophil function (including phagocytic activity, killing capacity, and oxidative burst activity) before initiation of therapy. The clinical significance of this finding is not clear.
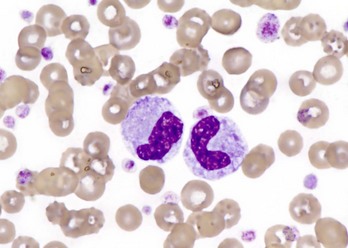
Fig. 13-20 Toxic change, neutrophils, canine blood smear.
Two band neutrophils with basophilic, foamy cytoplasm indicate toxic change. This dog also has reactive thrombocytosis. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Platelet function disorders, also known as thrombopathies or thrombopathias, may be primary or secondary. Many conditions are known or suspected to cause secondary platelet dysfunction (hypofunction or hyperfunction) by altering platelet adhesion, aggregation, or by mechanisms that are not fully understood. Box 13-3 shows underlying conditions associated with secondary platelet dysfunction.
Methods for Gross and Microscopic Examination
Routine Tests: Routine diagnostic testing often provides great insight into disorders of the hematopoietic system. The CBC is a cornerstone of diagnosis, part of routine evaluation of any sick patient. The CBC includes not only numeric data indicating the concentration of different cell types and hemoglobin in the blood but also information gleaned from microscopic review of a blood smear such as cell morphology and the presence of hemoparasites. (note: Some parasites may infect blood cells, such as Hepatozoon organisms within circulating neutrophils or monocytes or Bartonella organisms within erythrocytes, but mainly cause disease in other body systems and are therefore not discussed in this chapter.) Learning to evaluate blood smears is a valuable skill for any practicing veterinarian. The CBC also typically reports plasma protein concentration as measured with a refractometer. It is important to remember that changes in hydration status and in the distribution of body fluids between the vascular and extravascular compartments affect the concentration of both cells and proteins in the blood.
Additional Tests
• Bone marrow examination (aspiration or core biopsy, preferably both, for reasons given here) is usually done to assess hematopoiesis when a patient has unexplained cytopenia(s) or atypical cells in circulation but also to look for neoplasia or occasionally, infection. Preferred sites for collecting bone marrow samples are in horses, the sternum; in cattle, the proximal ribs; and in dogs and cats, the proximal humerus, iliac crest, or proximal femur. Normal hematopoietic cellularity of the marrow is age-dependent (recall that the amount of fat increases with age). Cytologic evaluation of a Wright’s-stained, air-dried smear of a bone marrow aspirate is the preferred method for evaluating cellular detail and in most cases provides a good estimate of overall marrow cellularity. Histopathologic evaluation of a hematoxylin and eosin (H&E) stained core biopsy is the most reliable way to assess marrow cellularity because tissue architecture is preserved. Bone marrow findings can only be interpreted fully along with a concurrent CBC. (note: Hematopoietic tissue is highly susceptible to autolysis. This is more of a limitation with regard to cytology than histology. Postmortem samples are often not suitable for cytologic interpretation unless collected immediately [i.e., within minutes after death].) Bone marrow examination yields information about the production of cells of specific lineages (i.e., erythropoiesis, granulopoiesis, and thrombopoiesis) in addition to the overall cellularity of the marrow. Increased production is evident as hyperplasia; decreased production as hypoplasia. The proportion of different cell types varies somewhat by species, but as a rule there are similar numbers of erythroid and myeloid (granulocytic and monocytic) nucleated cells, and much higher numbers of late-stage than early-stage cells. The amount of iron in the marrow can also be estimated by microscopic examination. Hemosiderin, a water-insoluble form of storage iron (less readily accessible than ferritin, which is soluble), is found mainly in macrophages in the marrow, spleen, and liver, and appears as coarse yellow-brown granular material with routine stains. Staining the sample with an iron-specific stain, such as Prussian blue, can make it easier to assess marrow iron stores (see Fig. 1-69). Cats differ from other species in that their marrow normally does not contain stainable iron.
• Aspiration cytology and/or histopathology of other organs (e.g., to assess extramedullary hematopoiesis, increased destruction of erythrocytes, neoplasia, or infection). Evaluation of lymph nodes and spleen is discussed later.
• Tests to detect an autoantibody bound to the cell surface. The Coombs’ test, or direct antiglobulin test, is the standard assay for IMHA. Flow cytometry and immunofluorescent antibody tests may also be used to detect autoantibody bound to erythrocytes or other cells.
• Immunophenotyping to identify cells based on expression of characteristic molecules (markers). Immunophenotyping may be done by immunohistochemistry, immunocytochemistry, or flow cytometry.
• Polymerase chain reaction (PCR) assays for infectious agents or clonal lymphocyte proliferations.
Hemostasis
Structural or functional abnormalities of blood vessels, platelets, or coagulation factors may result in a tendency toward hypocoagulability (bleeding), hypercoagulability (inappropriate thrombosis), or both. In veterinary medicine, there has been a great deal of work on specific mechanisms of hypocoagulability, whereas mechanisms of hypercoagulability are less fully characterized. Disorders of primary hemostasis typically result in “small bleeds” (e.g., petechiation, mild ecchymosis, bleeding from mucous membranes, bleeding immediately after venipuncture), whereas disorders of secondary hemostasis typically result in “big bleeds” (e.g., hemorrhage into body cavities/joints, marked ecchymosis, large hematomas, delayed bleeding after venipuncture). This chapter concentrates on primary disorders of hemostasis and also covers DIC, which is the secondary condition. However, it is important to note that coagulation disorders can also result from other underlying disease processes. For example, advanced liver disease can lead to abnormal hemostasis through decreased or defective synthesis of coagulation factors, or impaired clearance of fibrinolytic products that inhibit coagulation reactions and platelet function. Vascular disorders may also result in a bleeding tendency because of abnormalities of endothelial function or collagen-platelet interactions. Specific diseases involving abnormal structure or function of hematopoietic or hemostatic elements are discussed later in this chapter.
The CBC provides basic information about platelets, including numeric values for platelet concentration and mean platelet volume (MPV), subjective assessment of platelet morphology (size, shape, and granularity), and a rough estimation of platelet numbers based on examination of a blood smear. Some laboratories measure reticulated platelets (platelets recently released from the bone marrow), although this test is mostly used in the research setting at present. Increased MPV and increased numbers of reticulated platelets tend to indicate increased thrombopoiesis. Bone marrow examination is indicated with any unexplained cytopenia, including thrombocytopenia, to evaluate production. Tests to evaluate the components of the hemostatic process are described and listed in Web Appendix 13-1.
Disorders of Domestic Animals
Aplastic Anemia (Aplastic Pancytopenia)
Aplastic anemia is a rare condition characterized by aplasia or severe hypoplasia of all hematopoietic lineages in the bone marrow and resulting cytopenias. The term is something of a misnomer because affected cells are not limited to the erythroid lineage (see the later discussion of pure red cell aplasia in the section on Immune-Mediated Disorders). Aplastic anemia is also known more accurately as aplastic pancytopenia. Destruction of hematopoietic stem cells or progenitor cells is recognized as a cause of the condition. Other proposed mechanisms include disruption of normal stem cell function because of mutation or perturbation of the hematopoietic microenvironment.
Many of the conditions reported to cause aplastic anemia in domestic animals do so only rarely, or idiosyncratically, and more frequently cause other hematologic or nonhematologic abnormalities. A partial list of reported causes of aplastic anemia in domestic animals includes the following:
• Antimicrobial agents (dogs, cats)
• Chemotherapeutic agents (dogs, cats)
• Phenylbutazone (horses, dogs)
• Bracken fern (cattle, sheep)
Aplastic anemia occurs in both acute and chronic forms. Most of the chemical causes result in acute disease. As discussed in the earlier section on mechanisms of disease, severe neutropenia typically develops within 1 week of an acute insult to the bone marrow, after the bone marrow storage pool is depleted, and severe thrombocytopenia in the second week. The development of anemia is more variable, depending in part on how rapidly the marrow recovers from the insult, and on the RBC life span of the particular species. Severe neutropenia and thrombocytopenia predispose affected individuals to infection and hemorrhage, respectively. In addition to aplasia, pathologic bone marrow findings in animals with aplastic anemia may include evidence of necrosis, degeneration of hematopoietic cells, and an increase in phagocytic macrophages. Fig. 13-21 shows bone marrow aspirates from a dog with pancytopenia from acute 5-fluorouracil toxicosis, before and during recovery.
Fig. 13-21 Aplastic anemia, canine bone marrow aspirate.
A, Bone marrow aspirate from a dog 8 days after ingestion of a toxic dose of 5-fluorouracil shows stromal cells but a lack of developing blood cells. B, Bone marrow aspirate from the same dog one week later, after resumption of hematopoiesis. Inset, Higher magnification of Fig. 13-22, B shows early- and late-stage erythroid and granulocytic precursors. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Congenital Disorders
Many inherited or presumably inherited disorders of blood cells have been recognized in domestic animals, including rare or sporadic cases and conditions that are of questionable clinical relevance. This section and the later sections covering species-specific disorders are not comprehensive but instead focus on the more common, well-characterized, or recently reported conditions.
Erythropoietic Porphyrias
Porphyrias are inherited defects of enzymes involved in the synthesis of porphyrins, precursors of hemoglobin, and other heme proteins. Porphyrias result in accumulation of toxic porphyrin compounds. Congenital erythropoietic porphyria, transmitted as an autosomal recessive trait, occurs in Holstein and shorthorn cattle and is characterized by red-brown discoloration of teeth, bones, and urine caused by accumulation of porphyrins (see Fig. 1-75). Because of the circulation of the photodynamic porphyrins in blood, these animals have lesions of photosensitization of the nonpigmented skin and hemolytic anemia. All affected tissues, including erythrocytes, exhibit fluorescence with ultraviolet light. The premature destruction of developing and mature erythrocytes is caused by the accumulation within these cells of excess porphyrins. Bovine erythropoietic protoporphyria is an inherited disorder of heme synthetase, a terminal enzyme of the heme synthetic pathway, resulting in the accumulation of protoporphyrins in tissues and erythrocytes. It is inherited as an autosomal recessive trait and is confined to Limousin or Limousin-cross cattle. Photosensitivity is the only clinical manifestation of the disease; there is no anemia and no discoloration of teeth and bone. A congenital porphyria described in Siamese and domestic short-haired cats resembles congenital erythropoietic porphyria in cattle. These cats have brown teeth, photosensitization, and hemolytic anemia.
Pyruvate Kinase Deficiency
Pyruvate kinase (PK) deficiency is an inherited autosomal recessive condition reported in many dog breeds and fewer cat breeds (Abyssinian, Somali, and domestic shorthair). The glycolytic isoenzyme that is deficient in erythrocytes of affected animals normally catalyzes the last ATP-generating reaction in glycolysis. Thus there is decreased production of ATP in PK-deficient individuals, which results in loss of normal membrane function and decreased erythrocyte lifespan. The disease is characterized by moderate to severe extravascular hemolytic anemia that is strongly regenerative. In dogs, PK deficiency typically also involves progressive iron overload, myelofibrosis, osteosclerosis, and cirrhosis. Dogs with PK deficiency usually die from complications of the disease between 1 and 5 years of age. In cats, PK deficiency is associated with milder hematologic abnormalities and is not associated with osteosclerosis; the prognosis is more favorable. Dogs with PK deficiency do not necessarily have the same genetic defect, so mutation-specific DNA-based assays are required. In contrast, a single DNA-based test is available to detect the common mutation affecting Abyssinian, Somali, and domestic shorthair cats.
Cytochrome b5 Reductase Deficiency
Deficiency of cytochrome b5 reductase (Cb5R, also known as methemoglobin reductase), the enzyme that catalyzes the reduction of methemoglobin (Fe3+) to hemoglobin (Fe2+), has been recognized in many dog breeds and in domestic shorthair cats. It is probably an autosomal recessive trait. Affected animals may have cyanotic mucous membranes or exercise intolerance but are not anemic, usually lack clinical signs of disease, and have normal life expectancies.
Glucose-6-Phosphate Dehydrogenase Deficiency
Deficiency of the glycolytic enzyme glucose-6-phosphate dehydrogenase (G6PD), a common X-linked disease in people, has been reported in an American saddle bred colt with eccentrocytosis and persistent hemolytic anemia and in a partially deficient male dog without anemia or clinical signs.
Leukocyte Adhesion Deficiency
LAD is an inherited fatal autosomal recessive condition characterized by deficiency of the common β2 chain (also known as CD18) of leukocyte integrins. Without normal expression of this adhesion molecule, leukocytes have severely impaired ability to migrate from the blood into tissues. As a result, animals with LAD are highly susceptible to infections and usually die at a young age. Infected foci are nonsuppurative because of the absence of infiltrating leukocytes. LAD is also characterized by very high concentrations of neutrophils in the blood. LAD has been recognized in Holstein cattle (known as bovine LAD [BLAD]) and Irish setter dogs (see Chapter 3).
Pelger-Huët Anomaly
Pelger-Huët anomaly (PHA) is a condition characterized by lack of normal segmentation of the nuclei of mature granulocytes. This condition results in granulocyte morphology similar to that of band neutrophils in an inflammatory left shift; however, in the absence of any other disease process, animals with PHA should not have clinical signs or other laboratory findings indicating inflammation. PHA cells can be distinguished from immature forms of otherwise normal band forms on the basis of their mature chromatin pattern (Fig. 13-22). PHA has been described in dogs, cats, horses, and rabbits, as well as in humans. Prevalence is higher in certain dog breeds. In Australian shepherds, the mode of inheritance is autosomal dominant with incomplete penetrance. Most cases of PHA are the heterozygous form and probably of no clinical significance. In these cases, the shape of most granulocyte nuclei resembles those of band and metamyelocyte forms. A rare homozygous form of PHA has also been reported in rabbits and cats. It is associated with accompanying skeletal abnormalities and stillbirths or early mortality in these species (there are also rare reports of homozygous PHA in humans, without associated skeletal abnormalities or shortened lifespan). In homozygous PHA cells, granulocyte nuclei are round or oval. An acquired, reversible condition mimicking PHA, known as pseudo–PHA, is occasionally noted in animals with infectious disease, neoplasia, or in connection with drug administration.
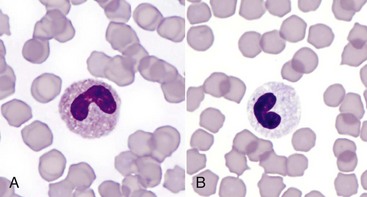
Chédiak-Higashi Syndrome
Chédiak-Higashi syndrome (CHS) is a rare autosomal recessive condition characterized by recurrent pyogenic infections, bleeding tendencies, ocular and cutaneous hypopigmentation, and prominent cytoplasmic inclusions in blood cells. CHS has been described in several breeds of cattle and in Persian cats. The mutated gene encodes a protein called beige or LYST (lysosome trafficking regulator), the specific functions of which are still being investigated. Individuals with CHS have severely impaired cellular innate immunity because of neutropenia, impaired chemotaxis, and impaired killing by granulocytes and cytotoxic lymphocytes.
One of the classic features of CHS is a bleeding tendency caused by platelet dysfunction. Platelets in individuals with CHS lack the dense granules that normally contain key bioactive molecules involved in hemostasis, including platelet agonists, such as ADP and serotonin, and platelet aggregation in vitro in response to collagen, in particular, is severely impaired.
Glanzmann Thrombasthenia
Glanzmann thrombasthenia (GT) is an inherited platelet function defect caused by defective expression of the integrin αIIbβ3 (also known as GPIIb-IIIa). The αIIbβ3 molecule has multiple functions but is best known as a fibrinogen receptor that is essential for normal platelet aggregation. Bleeding tendencies vary widely between affected individuals, and bleeding tends to occur mainly on mucosal surfaces. The condition is characterized by an in vitro lack of response to all platelet agonists and severely impaired clot retraction. GT has been recognized in Great Pyrenees and Otterhound dogs and in a quarter horse, a thoroughbred-cross, and an Oldenburg filly. To date, all reported cases in animals have involved mutations to the IIb gene. Molecular testing is available to detect affected or carrier Great Pyrenees and Otterhound dogs and to detect the reported mutations in horses.
CalDAG-GEFI Thrombopathia
Conditions characterized by inability to relay signaling information in response to most platelet agonists have been described in basset hound, Eskimo spitz, and Landseer-ECT dogs and in Simmental cattle. The molecular basis involves mutations to the gene encoding the guanine nucleotide exchange factor, CalDAG-GEFI. Platelet receptor expression in affected animals is apparently normal. All reported mutations have been associated with a bleeding tendency.
Toxicoses
Rather than presenting an exhaustive list, this section describes some of the more commonly recognized oxidants that cause hemolytic anemia or impaired hemoglobin function in common domestic species. Oxidative insult may result in extravascular hemolysis because of phagocytosis of damaged erythrocytes by splenic macrophages, or in intravascular hemolysis if the damage is severe enough. In horses, red maple (Acer rubrum) toxicity is a well-characterized, potentially fatal cause of acute intravascular hemolytic anemia and methemoglobinemia. Ingestion of sufficient amounts of wilted or dried leaves or bark causes Heinz body formation, eccentrocytosis, severe intravascular hemolysis, and methemoglobinemia. Common postmortem findings include icterus, splenic hemosiderosis, splenomegaly, brown discoloration and swollen liver, and swollen kidneys that can be dark red to blue-black. Histologically, the kidneys have characteristic red-brown tubular casts (hemoglobinuric nephrosis).
Phenothiazine can cause Heinz body formation in horses. In ruminants, Brassica and rye grass are associated with Heinz body formation, and nitrite toxicity causes methemoglobinemia.
Copper toxicity is a well-known cause of acute intravascular hemolytic anemia in sheep and may also occur in goats and calves. The condition occurs in animals that have accumulated large amounts of copper in the liver. The copper is released under conditions of stress (e.g., shipping, starvation) and is believed to cause hemolysis as a result of direct interaction with membrane proteins, lipid peroxidation, formation of reactive oxygen species, and enzyme inhibition. Affected animals are often markedly icteric, and hemoglobinuric nephrosis is a classic postmortem lesion.
Onions and garlic are most commonly recognized as a cause of Heinz body hemolytic anemia in dogs and cats but can cause oxidative damage to erythrocytes in other domestic animals, including horses and cattle. Other causes of Heinz bodies in dogs include acetaminophen, benzocaine, naphthalene, phenylhydrazine, vitamins K1 and K3, and zinc toxicity. Other causes of Heinz bodies in cats include acetaminophen, benzocaine, methionine, naphthalene, propofol, and propylene glycol.
Nutritional and Metabolic Disorders
Severe malnutrition is probably a cause of nonregenerative anemia in all species attributable to combined deficiencies of molecular building blocks, energy, and essential cofactors. By far the most commonly recognized specific deficiency that results in anemia is iron deficiency. Iron deficiency is usually not a primary nutritional deficiency but rather occurs secondary to depletion of iron stores via chronic hemorrhage. The most common route of loss is through the GI tract (e.g., a result of neoplasia, especially GI carcinoma or lymphoma, in older animals or hookworm infection in puppies). Chronic hemorrhage may also be caused by many other things, including marked ectoparasitism (e.g., pediculosis in cattle and massive flea burden in kittens and puppies), neoplasia in locations other than the GI tract, and bleeding diatheses. Rapidly growing nursing animals may be iron-deficient when compared with adults because milk is an iron-poor diet. In most cases, this has little clinical significance (and in fact is normal). An important exception is piglets with no access to iron, which may cause anemia, failure to thrive, and increased mortality. Neonatal piglets are routinely given parenteral iron (intramuscular injection of iron dextran) for this reason.
Other specific nutritional deficiencies causing anemia in animals are uncommon or rare. Copper deficiency can cause iron deficiency in ruminants, and may occur because of copper-deficient forage or impaired usage of copper by high dietary molybdenum or sulfate. It is believed that copper deficiency impairs production of ceruloplasmin, a copper-containing enzyme involved in GI iron absorption. Copper deficiency anemia, like iron deficiency anemia, is typically microcytic and hypochromic. Cobalamin (vitamin B12) and folate deficiencies are recognized as causes of anemia in humans but are rare in animals.
Infectious Diseases
This section covers infectious agents within the same genus that are recognized to cause disease in multiple species. Other infectious agents with more limited host specificity (for example, cytauxzoonosis in cats, feline and equine retroviruses) are covered in later sections on species-specific diseases. Throughout both sections, diseases are organized by taxonomy (protozoal, bacterial and rickettsial, and viral).
Babesiosis
Babesia spp. are intraerythrocytic protozoal organisms (piroplasmas) spread by arthropods (ticks, biting flies), transplacentally, and by blood transfusions. Evidence is accumulating that infection also is transmitted by dog fighting. Babesia spp. cause hemolytic anemia in horses, cattle, dogs, cats, and various nondomesticated animals. Babesia organisms are typically classified as large or small form, based on routine light microscopic morphology. Examples of large and small form organisms are shown in Fig. 13-23. Babesia equi and B. caballi infect horses and other equids in tropical and subtropical areas worldwide (B. equi, which also infects lymphocytes, is not considered a true Babesia but is phylogenetically more closely related to Theileria and Cytauxzoon). B. bovis and B. bigemina (small and large form organisms, respectively) are pathogenic Babesia spp. in cattle. These organisms have a worldwide distribution but have been eradicated in North America. Other, less pathogenic Babesia spp. may also infect cattle. In dogs, large form and small form Babesia spp. infections are associated with hemolytic anemia. The predominant large form is B. canis, which has three subtypes (canis, rossi, and vogeli). B. canis vogeli, considered the least pathogenic strain, is the most common one in the United States (US). In some areas, particularly the southeastern US, the seroprevalence of B. canis is high, and many infected dogs are asymptomatic chronic carriers. There are also reports of dogs with other types of large form babesiosis, including one caused by an organism with highest molecular homology to B. bovis. Small form Babesia spp. in dogs include B. gibsoni, B. conradae, and a form known as the Spanish isolate (formerly Theileria annae). A number of Babesia spp. (B. cati, B. felis, B. herpailuri, B. pantherae, and others) have been reported in domestic and wild cats worldwide. Little is known about the epidemiology of feline babesiosis; the severity of disease ranges from asymptomatic to severe.
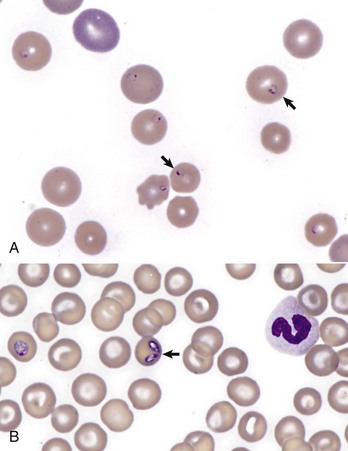
Fig. 13-23 Babesiosis, canine blood smear.
A, Small form (arrows) of Babesia (consistent with Babesia gibsoni). B, Babesia canis (arrow) organisms infecting erythrocytes. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Babesiosis may cause both intravascular and extravascular hemolysis and is also associated with a wide range of other clinical signs. The wide variety of clinical signs is a result of variations in pathogenicity of the organisms and susceptibility of the host. Infection with highly virulent strains may cause severe multisystemic disease. In these cases, massive immunostimulation and cytokine release cause circulatory disturbances, which may result in shock, induction of the systemic inflammatory response, and multiple organ dysfunction syndromes. Mechanisms of hemolysis may include direct damage to the erythrocyte by protozoal proteases, immune-mediated destruction, and oxidative damage. In animals with acute disease, signs often include fever, lethargy, pallor, hemoglobinuria, splenomegaly, and icterus. Animals are often thrombocytopenic, presumably a result of immune-mediated destruction of platelets. They also may have lymphadenopathy. Less common signs include edema, ascites, signs of central nervous system dysfunction, renal failure, rhabdomyolysis, stomatitis, and gastroenteritis.
Babesia organisms can usually be detected on a routine blood smear in animals with acute disease. Infected erythrocytes may be more prevalent in capillary blood, so blood smears made from samples taken from the pinna of the ear or the nail bed may increase the likelihood of detecting organisms microscopically. Buffy coat smears also have an enriched population of infected RBCs. PCR testing is the most sensitive assay for detecting infection in animals with very low levels of parasitemia. At necropsy, animals dying of the acute disease have splenomegaly, jaundice, hemoglobinuria, swollen hemoglobin-stained kidneys, and subepicardial and subendocardial hemorrhages. The cut surface of the enlarged, congested spleen oozes blood. The gallbladder is usually distended with thick bile. A striking feature of B. bovis infections is congestion of gray matter throughout the brain, which is readily visible compared with the white matter. Parasitized erythrocytes are best visualized on impression smears of the kidney, brain, and skeletal muscle. Microscopic findings in the liver and kidney are typical of a hemolytic crisis and include anemia-induced degeneration, necrosis of periacinar hepatocytes and cholestasis, and hemoglobinuric nephrosis with degeneration of tubular epithelium. Erythroid hyperplasia is present in the bone marrow. In animals that survive the acute disease, there is hemosiderin accumulation in the liver, kidney, spleen, and bone marrow. In chronic cases, there is hyperplasia of macrophages in the red pulp of the spleen.
Theileriosis
Theileria spp. are tick-borne protozoal organisms that infect many domestic and wild artiodactyls worldwide. Infection is characterized by intralymphocytic schizonts and pleomorphic intraerythrocytic piroplasmas (merozoites and trophozoites). The latter stages closely resemble Cytauxzoon and small form Babesia spp. Recognized Theileria pathogens in cattle include Theileria parva (the cause of East Coast fever in Africa), T. annulata (the cause of tropical theileriosis in the Mediterranean regions, Middle East, and Asia), and T. buffeli, which has recently been documented as a cause of hemolytic anemia in the US. Possible mechanisms of anemia in theileriosis include invasion of erythroid precursors by merozoite stages and associated erythroid hypoplasia (as occurs with T. parva infection), immune-mediated hemolysis, mechanical fragmentation because of vasculitis or microthrombi, enzymatic destruction by proteases, and oxidative damage. Clinical signs in a severely anemic cow infected with T. buffeli included recumbency, fever, pallor, tachycardia, and lymphadenopathy. Necropsy findings included splenic hemosiderosis, edema of lymph nodes and the subcutis, thoracic and peritoneal effusions, and pneumonia.
Trypanosomiasis
Trypanosomes are flagellated protozoa that normally survive and are nonpathogenic in wildlife reservoir hosts. They are transmitted by arthropod vectors. Some Trypanosoma spp. are recognized as causing hemolytic anemia in animals in tropical and subtropical regions outside of North America. Trypanosoma brucei and T. evansi affect horses. T. congolense and T. vivax affect cattle. Trypanosoma spp. that cause anemia in other species include T. simiae in pigs and T. brucei in camels and horses. Trypanosomes also cause nonhemolytic disease (e.g., T. cruzi, the agent of Chagas’ disease, or American trypanosomiasis) in many hosts, and nonpathogenic variants (e.g., T. theileri in cattle worldwide) are also recognized. Trypanosomal organisms do not infect erythrocytes but rather exist as free trypomastigotes (flagellated forms with a characteristic undulating membrane) in the blood (Fig. 13-24, A) or as amastigotes in tissue. The mechanism of anemia is believed to be immune-mediated. Cattle with acute trypanosomiasis have significant anemia, which initially is regenerative, but less so with time. The extent of parasitemia is readily apparent with T. vivax and T. theileri infections because the organisms are present in large numbers in the blood. This is in contrast to T. congolense, which localizes within the vasculature of the brain and skeletal muscle. Cattle with T. congolense infections develop chronic debilitating disease; they have scruffy hair coats, appear “potbellied,” and have fever, intermittent diarrhea, and exercise intolerance. Mortality is greater with T. vivax infections, usually because of intercurrent acute infectious diseases such as salmonellosis. Necropsy findings in cattle with trypanosomiasis include cachexia, generalized edema with increased fluid in body cavities, and generalized lymph node enlargement. Bronchopneumonia, a flabby heart, and serous atrophy of pericardial fat may be present. Liver and kidneys are symmetrically enlarged. Lymph nodes are enlarged up to four times normal size because of lymphoid hyperplasia, and most of the fatty bone marrow is replaced by red hematopoietic tissue. The spleen is enlarged because of lymphoid hyperplasia and is firm when incised.
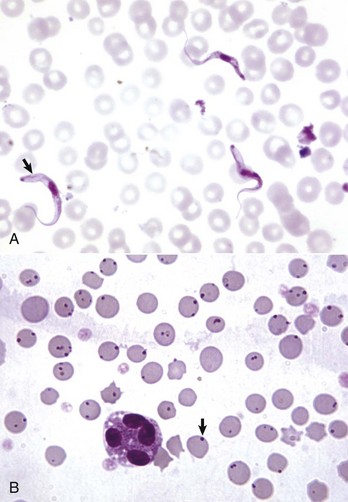
Fig. 13-24 Hemotropic parasites, bovine blood smear.
A, Trypanosomiasis. Trypanosomes are flagellated protozoa (arrow) characterized by an undulating membrane, kinetoplast, and nucleus. They may be identified in a wet mount made from the buffy coat portion of the packed cells. B, Anaplasmosis, Anaplasma marginale. Note the darkly stained organisms (arrow), most of which are located on the edges of the erythrocytes. Anaplasmosis causes anemia mainly by immune-mediated extravascular hemolysis. (A courtesy Dr. M.D. McGavin, College of Veterinary Medicine, University of Tennessee. B courtesy Dr. J. Simon, College of Veterinary Medicine, University of Illinois.)
Anaplasmosis and Ehrlichiosis
Anaplasma spp. are rickettsial organisms that may be transmitted by arthropod vectors (ticks, biting flies) or by blood-contaminated needles. Anaplasmosis is a cause of hemolytic anemia in cattle in tropical and subtropical areas of many parts of the world. Anaplasma marginale, considered the more pathogenic species, has a worldwide distribution. A. centrale is found in South America, Africa, and the Middle East. The genus names reflect the typical locations of the organisms when detected on examination of a blood smear, either on the periphery or more centrally within infected erythrocytes. A related species, A. ovis, affects sheep and goats in tropical and subtropical areas worldwide. Wild animals, such as deer, elk, and bison, may be latently infected and serve as reservoir hosts for A. marginale. Anaplasma organisms infect erythrocytes intracellularly.
Anaplasmosis causes anemia mainly by immune-mediated extravascular hemolysis. The severity of disease in infected animals varies with age. Infected calves under 1 year of age rarely develop clinical disease, whereas cattle 3 years of age or older are more likely to develop severe, potentially fatal, illness. The reason for this discrepancy is not clear. In clinically affected animals, common signs include lethargy or recumbency, pallor, and icterus. Animals dying of acute anaplasmosis have blood of low viscosity, pale to icteric tissue, an enlarged turgid spleen, and an icteric liver with a distended gallbladder. In animals with acute disease, it is usually easy to detect A. marginale organisms on routine blood smear evaluation (Fig. 13-24, B). However, in recovering animals, the organisms may be difficult to find. Surviving cattle become chronic carriers (and thus reservoirs for infection of other animals) and develop cyclic parasitemia, which is typically not detectable on blood smears. Splenectomy of carrier animals results in marked parasitemia and acute hemolysis. PCR testing is the most sensitive means of identifying animals with low levels of parasitemia.
Ehrlichiae are small, pleomorphic, Gram-negative, obligate intracellular bacteria that are transmitted by tick vectors. Some Ehrlichia spp. have a tropism for granulocytes, and morulae are sometimes found within the cytoplasm of neutrophils of affected animals (Fig. 13-25). Molecular analysis has shown that organisms previously considered to be distinct entities—E. equi, the agent of human granulocytic ehrlichiosis, and E. phagocytophilum—are genetically indistinguishable. All are now designated as Anaplasma phagocytophilum. Granulocytic ehrlichiosis occurs naturally in horses (A. phagocytophilum), dogs (A. phagocytophilum and E. ewingii), and cats (A. phagocytophilum). A. phagocytophilum survives within neutrophils by dysregulating key bactericidal functions, including the NADPH oxidase system, and delaying neutrophil apoptosis. Common clinical manifestations of infection include fever and signs referable to processes affecting cell or tissue types other than neutrophils (e.g., thrombocytopenia, anemia, immune-mediated polyarthritis). Neutropenia may occur in animals infected with A. phagocytophilum, but the mechanism is not clear. Judging from serologic evidence of exposure to A. phagocytophilum, most naturally infected dogs probably stay healthy.
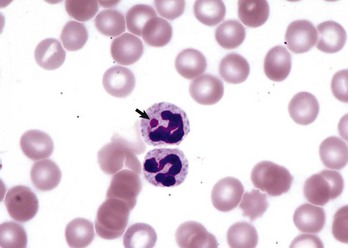
Fig. 13-25 Granulocytic ehrlichiosis, Anaplasma phagocytophila, canine blood smear.
The top neutrophil contains an inclusion (arrow) consistent with an Anaplasma phagocytophila morula. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Some Ehrlichieae have a tropism for mononuclear cells, but clinical manifestations are usually referable to other cell types or body systems. Morulae may be found in mononuclear cells of infected animals on routine examination of blood smears, and examination of buffy coat smears increases the probability of detecting the organism. In horses, E. risticii—the agent of Potomac horse fever (also known as equine monocytic ehrlichiosis)—infects monocytes and enterocytes and is primarily a diarrheal disease (see Chapter 7). In dogs, E. canis, the agent of canine monocytic ehrlichiosis, infects mononuclear cells. Infection with E. chaffeensis, the agent of human monocytic ehrlichiosis, has also been reported in dogs. Canine monocytic ehrlichiosis has acute and chronic forms. Acutely ill animals typically have a fever, lymph node enlargement, and splenomegaly. Thrombocytopenia and nonregenerative anemia are common findings. Untreated dogs recovering from acute disease develop a subclinical phase and may have persistent mild thrombocytopenia. A subset of these dogs develop chronic disease that may be debilitating and in some cases life threatening. Some studies indicate that German shepherd dogs with ehrlichiosis are predisposed to have particularly severe clinical disease. Severe cases are characterized by weight loss, lymph node enlargement, pyrexia, thrombocytopenia, and nonregenerative anemia. Thrombocytopenic animals may have severe bleeding tendencies. Some dogs with chronic disease develop pancytopenia. Necropsy findings vary with the stage of the disease. In the acute disease, there are widespread petechiae and ecchymoses, with splenomegaly and lymphadenomegaly. Chronically infected dogs are emaciated. The bone marrow is hyperplastic and red in the acute disease, but becomes hypoplastic and pale in animals with pancytopenia. Histologic findings include generalized perivascular plasma cell infiltration, which is most pronounced in animals with chronic disease. Multifocal, nonsuppurative meningoencephalitis, interstitial pneumonia, and glomerulonephritis are present in most dogs with the disease. Ehrlichia organisms are difficult to detect histologically; examination of Wright-Giemsa–stained impression smears of lung, liver, lymph nodes, and spleen is a more effective method for detecting the morulae in macrophages. Ehrlichiosis is often diagnosed on the basis of serologic testing, but PCR testing is more sensitive.
Anaplasma platys (previously called Ehrlichia platys) is a rickettsial organism that infects canine platelets, causing recurrent marked thrombocytopenia (the disease is also known as infectious canine cyclic thrombocytopenia). The disease is tick-borne and has been recognized worldwide. Evidence of megakaryocytic hyperplasia and organism-associated antigen within macrophages indicate that thrombocytopenia likely results from increased platelet destruction. Infection is generally considered to be asymptomatic, and morulae within platelets may be detected on blood smears incidentally, but there are rare reports describing more severe clinical signs in infected animals.
Clostridial Diseases
Certain Clostridium spp. may cause potentially fatal hemolytic anemias in animals. The mechanism of hemolysis involves a bacterial toxin (phospholipase C or lecithinase), which enzymatically degrades cell membranes, causing acute intravascular hemolysis. C. haemolyticum and C. novyi type D cause the disease in cattle known as bacillary hemoglobinuria. (The phrase “red water” has also been used for this disease and for hemolytic anemias in cattle caused by Babesia spp.) Similar naturally occurring disease has been reported in sheep and elk. In cattle, the disease is associated with liver fluke (Fasciola hepatica) migration in susceptible animals. Ingested clostridial spores may live in Kupffer cells of the liver for a long time without causing disease. However, when migrating flukes cause hepatic necrosis, the resulting anaerobic environment stimulates the clostridial organisms to proliferate and elaborate their hemolytic toxins, causing additional hepatic necrosis. Bacillary hemoglobinuria has also been associated with liver biopsy in calves. C. perfringens type A causes intravascular hemolytic anemia in lambs and calves—a condition known as yellow lamb disease, yellows, or enterotoxemic jaundice because of the characteristic jaundice. The organism is a normal inhabitant of the GI tract in these animals, but may proliferate abnormally in response to some diets. C. perfringens has also been associated with intravascular hemolytic anemia in horses with clostridial abscesses and in a ewe with clostridial mastitis.
Leptospirosis
Leptospirosis is recognized as a cause of hemolytic anemia in calves, lambs, pigs, and black rhinoceros. Specific leptospiral organisms associated with hemolytic disease include Leptospira interrogans serovars pomona and ictohaemorrhagiae. Proposed mechanisms of disease include immune-mediated (IgM cold agglutinin) extravascular hemolysis and enzymatic (phospholipase produced by the organism) intravascular hemolysis.
Leptospira organisms are ubiquitous in the environment. Infection occurs percutaneously and via mucosal surfaces and is followed by leptospiremia; organisms then localize preferentially in certain tissues (e.g., kidney, liver, and pregnant uterus). Leptospirosis can also cause many disease manifestations besides hemolysis (e.g., renal failure, liver failure, abortion, and other conditions) that are not discussed here.
In addition to anemia, common findings in animals with leptospirosis-induced hemolysis include hemoglobinuria and icterus. On necropsy, renal tubular necrosis, which occurs in part because of hemoglobinuria (hemoglobinuric nephrosis), may also be present.
Hemotropic Mycoplasmosis
The term hemotropic mycoplasmas, or hemoplasmas, encompasses a group of bacteria, formerly known as Haemobartonella or Eperythrozoon spp., that commonly infect RBCs of many domestic, laboratory, and wild animals. Hemotropic mycoplasmas affecting common domestic species are listed in Table 13-2. The mode of transmission is poorly understood but arthropods are believed to play a role; transmission in utero and through biting or fighting is also suspected. Effects of infection vary from subclinical to fatal anemia, depending on the specific organism, dose, and host susceptibility. Anemia occurs mainly because of extravascular hemolysis. Although the pathogenetic mechanisms are not completely understood, an immune-mediated component is highly probable. Hemotropic mycoplasmas (and nonhemotropic Mycoplasma spp.) induce cold agglutinins in infected individuals, although it is not clear whether these particular antibodies are important in the development of hemolytic anemia. Like other mycoplasmas, hemotropic mycoplasmas are small (0.3 to 3 µm in diameter), are Gram-negative, and lack a cell wall. They are epicellular parasites, residing in indentations and invaginations of the RBC surface. When detected on routine blood smear evaluation, the organisms are variably shaped (cocci, small rods, or ring forms) and sometimes arranged in short, branching chains (especially M. haemocanis). The organisms may also be noted extracellularly, in the background of the blood smear, especially if the smear is made after prolonged storage of the blood in an anticoagulant tube. A photomicrograph of hemotropic mycoplasma organisms in a peripheral blood smear are shown in Fig. 13-26.
TABLE 13-2
Common Hemotropic Mycoplasmas in Domestic Animals
| Host | Organism |
| Cattle | Mycoplasma wenyonii (formerly Eperythrozoon wenyonii) |
| Dog | Mycoplasma haemocanis (formerly Haemobartonella canis) |
| Cat | Mycoplasma haemofelis (formerly Haemobartonella felis large form or Ohio variant) |
| Cat | Candidatus Mycoplasma haemominutum (formerly Haemobartonella felis small form or California variant) |
| Cat | Candidatus Mycoplasma turicensis (to date, has only been identified by molecular methods) |
| Pig | Mycoplasma suis (formerly Eperythrozoon suis) |
| Alpaca | Candidatus Mycoplasma haemolamae |
Fig. 13-26 Hemotropic mycoplasmosis, porcine blood smear.
Blood smear from a splenectomized pig infected with Mycoplasma suis (formerly Eperythrozoon suis). Note the small oval- to ring-shaped organisms attached to the surface of the erythrocytes and free in the protein of the blood smear. Wright’s stain. (Courtesy Dr. J.F. Zachary, College of Veterinary Medicine, University of Illinois.)
Most hemotropic mycoplasma subspecies are more likely to cause acute illness in individuals that are immunocompromised or have concurrent disease. M. haemofelis infection is an exception, causing acute hemolytic anemia in immunocompetent cats. The disease in sheep and pigs has a seasonal incidence corresponding to the peak occurrence of biting insects. However, it can also occur at other times of the year as a recrudescence in a carrier animal secondary to another disease. In both sheep and pigs, unexpected death of one or two animals is often followed by anemia in other animals within the herd. M. wenyonii in cattle is less pathogenic than M. ovis and M. suis in sheep and pigs, respectively. Infection with M. wenyonii appears to be widespread, but it rarely causes disease. Clinical signs in animals with acute disease include lethargy, fever, and pallor. Affected animals usually have mild to moderate hyperbilirubinemia and may be icteric. Animals probably remain chronically infected after recovery, even if treated with appropriate antibiotics. Chronically infected cattle, sheep, and pigs may have decreased production. Chronically infected dogs and cats are typically asymptomatic. In dogs, M. haemocanis infection is usually subclinical in immunocompetent animals but causes acute hemolytic anemia when infected animals receive a splenectomy. Two forms of hemoplasmas are known to infect cats. As mentioned earlier, M. haemofelis, the large form variant, causes acute hemolytic anemia in immunocompetent animals. Cats infected with the small form variant, which has the proposed name M. haemominutum, are typically asymptomatic or have only mild disease. Organisms are often but not always detected on routine blood smear examination in acutely ill animals. PCR testing is the most sensitive means of detecting infection in animals with low levels of parasitemia. In animals dying of hemotropic mycoplasma infection, the findings are typical of extravascular hemolysis, with pallor, icterus, and splenomegaly, and distended gallbladder (Fig. 13-27). Microscopic lesions in the spleen include congestion, erythrophagia, macrophage hyperplasia, extramedullary hematopoiesis, and increased numbers of plasma cells. Bone marrow has varying degrees of erythroid hyperplasia, depending on the duration of hemolysis.
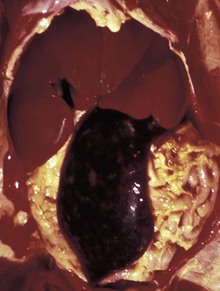
Fig. 13-27 Mycoplasma haemofelis, cat.
Note the splenomegaly, hepatomegaly, and icterus caused by infection of erythrocytes with this hemotropic parasite. Splenomegaly and icterus are the result of increased destruction (extravascular hemolysis) of infected erythrocytes. (Courtesy College of Veterinary Medicine, University of Illinois.)
Immune-Mediated Disorders
Immune-Mediated Hemolytic Anemia
IMHA is a condition characterized by increased destruction of erythrocytes because of binding of immunoglobulin to cell surface antigens. It is a common, life-threatening condition in dogs and also has been described in horses, cattle, and cats. Although the clinical picture of IMHA is variable, it typically has an acute onset and causes severe anemia. Some studies have shown that certain dog breeds (cocker spaniels and others) are predisposed to develop IMHA, suggesting the possibility of a genetic component, and the disease is more common in young to middle-aged female dogs. In most cases the reactive antibody is IgG, and hemolysis is extravascular (i.e., RBCs with bound antibody are phagocytosed by macrophages, mainly in the spleen). IgM and/or complement proteins may also contribute to IMHA. Complement usually acts as an opsonin (C3b) that promotes phagocytosis. However, formation of the complement membrane attack complex and resulting intravascular hemolysis is also a recognized mechanism and more likely to occur with IgM autoantibodies. Most immunoglobulins implicated in IMHA are reactive at body temperature (warm hemagglutinins). A smaller portion, usually IgM, are more reactive at lower temperatures and may lead to a condition known as cold hemagglutinin disease—ischemic necrosis at anatomic extremities (e.g., tips of the ears), where cooling of the circulation causes autoagglutination of erythrocytes and occlusion of the microvasculature. Typically, IMHA targets mature erythrocytes and is accompanied by a marked regenerative response. However, as discussed earlier in the chapter, immune-mediated destruction of immature erythroid cells in the bone marrow may also occur, resulting in nonregenerative anemia.
In veterinary medicine, IMHA is usually idiopathic (also called primary IMHA or autoimmune hemolytic anemia), and the specific trigger for the autoimmune reaction is not clear. Factors implicated in secondary IMHA include infection, drug administration, vaccination, neoplasia, and bee sting envenomation. Diagnosis of secondary IMHA is most often based on circumstantial evidence and exclusion of other known causes of hemolytic anemia, rather than on controlled experiments proving a direct causal relationship. Infectious agents affecting blood cells are discussed in more detail later. Drugs or chemicals associated with or suspected of causing IMHA in animals include antibiotics (cephalosporins, penicillin, and sulfonamides), levamisole, propylthiouracil, and the insecticide pirimicarb. Most drug-induced IMHA is believed to occur because of drug or drug metabolite interacting with the erythrocyte plasma membrane. Other proposed mechanisms include binding of drug-antibody immune complexes to the erythrocyte membrane, or induction of a true autoantibody directed against an erythrocyte antigen. A case of suspected vaccine-associated IMHA has been reported in a cow. Certain vaccines used in cows have been incriminated in the development of a specific form of IMHA in newborn calves: neonatal isoerythrolysis, which is discussed later. Retrospective studies investigating the relationship between vaccination history and development of IMHA in dogs have yielded conflicting results and are inconclusive.
Other common clinical findings in patients with IMHA include hyperbilirubinemia and splenomegaly (see Fig. 13-9), pyrexia, and inflammatory neutrophilia. These abnormalities vary in magnitude depending on the severity and duration of disease. Dogs with IMHA are also predisposed to develop hemostatic abnormalities (prolonged coagulation times, decreased plasma antithrombin [AT] concentration, increased plasma concentration of FDPs/D-dimer, thrombocytopenia, and DIC). The severity of postmortem lesions in dogs with IMHA, including ischemic necrosis within vital organs (liver, kidney, heart, and lung) and the spleen as a result of thromboembolic disease or hypoxia, has been shown to correlate with the severity of leukocytosis. Intravascular hemolysis (IVH) plays a relatively insignificant role in most cases of IMHA, but evidence of IVH (hemoglobinemia, hemoglobinuria) is noted occasionally, presumably in those cases in which IgM and complement are major mediators of hemolysis.
Neonatal Isoerythrolysis
A form of IMHA whose specific pathogenesis is well understood is neonatal isoerythrolysis (NI), a condition in which a newborn has colostrum-derived maternal antibodies, which react against its own erythrocytes. NI is common in horses (Fig. 13-28) and has been reported in cattle, cats, and some other domestic and wildlife species. In horses, this situation occurs as a result of immunosensitization of the dam from exposure to an incompatible blood type inherited from the stallion (e.g., transplacental exposure to fetal blood during pregnancy or mixing of maternal and fetal blood during parturition). A previously mismatched blood transfusion produces the same results. Some equine blood groups are more antigenic than others; in particular, types Aa and Qa are very immunogenic in mares. Severely affected foals become lethargic and weak as early as 8 to 10 hours after birth or at any time during the succeeding 4 to 5 days. They have pale, icteric mucous membranes and may have hemoglobinuria. Serum bilirubin concentrations are usually increased, and foals that die during a hemolytic crisis are notably icteric and have splenomegaly. In cattle, NI has been associated with vaccination with whole blood products or products containing erythrocyte membrane fragments. In cats, the recognized form of NI does not depend on prior maternal immunosensitization but on naturally occurring anti-A antibodies in queens with Type B blood. NI has been produced experimentally in dogs, but there are no reports of naturally occurring disease. NI can be prevented by typing the maternal and paternal blood and not allowing neonates from incompatible matings access to the mother’s colostrum, or by not allowing animals with strongly incompatible blood types to breed.
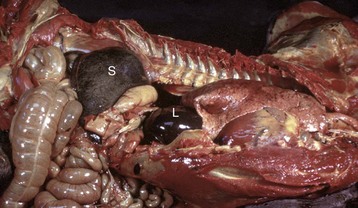
Fig. 13-28 Neonatal isoerythrolysis, foal.
Note the enlarged spleen (S) (also liver [L]) and icterus. The newborn foal had colostrum-derived maternal antibodies, which reacted against its own erythrocytes. Macrophages in the splenic red pulp remove erythrocytes whose membranes have bound antibody. (Courtesy College of Veterinary Medicine, University of Illinois.)
Pure Red Cell Aplasia
Pure red cell aplasia (PRCA) is a rare bone marrow disorder characterized by absence of erythropoiesis and severe nonregenerative anemia. Primary and secondary forms of PRCA have been described in dogs and cats. Primary PRCA is apparently caused by immune-mediated destruction of early erythroid progenitor cells, a presumption supported by the response of some patients to immunosuppressive therapy and by the detection of antibodies inhibiting erythroid colony formation in vitro in some dogs. Infection with FeLV subgroup C is associated with secondary erythroid aplasia in cats, probably because of infection of early stage erythroid precursors. Parvoviral infection has been suggested as a possible cause of secondary PRCA in dogs. Administration of recombinant human Epo (rhEpo) has been identified as a cause of secondary PRCA in dogs, cats, and horses, presumably caused by induction of antibodies against rhEpo that cross-react with endogenous Epo. Experimentation with the use of species-specific recombinant Epo has produced mixed results. Dogs treated with recombinant canine Epo have not developed PRCA. However, in experiments reported thus far involving cats treated with recombinant feline Epo, at least some animals have developed PRCA.
Immune-Mediated Neutropenia
Immune-mediated neutropenia is a rare condition that has been reported in horses, dogs, and cats. The range of etiologies is presumably similar to that of the better characterized immune-mediated cytopenias (anemia and thrombocytopenia). The diagnosis may be supported by flow cytometric detection of immunoglobulin bound to neutrophils but is most often made on the basis of exclusion of other causes of neutropenia and response to immunosuppressive therapy.
Immune-Mediated Thrombocytopenia
Immune-mediated thrombocytopenia (IMT) is a condition characterized by immune-mediated destruction of platelets. There are numerous similarities between IMT and IMHA. IMT is a fairly common condition in dogs (it has also been described in horses and cats). It is more common in middle-aged animals, females, and perhaps some breeds of dogs. The disease is usually idiopathic and typically results in severe thrombocytopenia (often <10,000 platelets/µL), although less severe forms of the disease also occur. Affected animals have varying degrees of clinical bleeding tendencies. IMT also sometimes occurs together with IMHA, a condition known as Evans syndrome.
Underlying conditions associated with secondary IMT include infection (e.g., equine infectious anemia virus [EIAV] and ehrlichiosis), drug administration, neoplasia, and other immune-mediated diseases. Many types of infectious agents (viral, bacterial, fungal, and protozoal) have been associated with IMT, some of which may also cause thrombocytopenia by other mechanisms. Ehrlichiosis, for example, has been shown to elicit production of antibodies that are cross-reactive with platelet antigens in humans. Drugs suspected of causing IMT in animals include cephalosporins and sulfonamides.
Neonatal Alloimmune Thrombocytopenia
A form of immune-mediated thrombocytopenia, known as neonatal alloimmune thrombocytopenia, is recognized in neonatal pigs and foals. The pathogenesis of this disease is virtually identical to that of neonatal isoerythrolysis as a cause of anemia: a neonate inheriting paternal platelet antigens absorbs maternal antibodies against these antigens through the colostrum. In principle, a similar situation may occur after platelet-incompatible transfusion of blood or blood products containing platelets.
Inflammatory Disorders
Hemophagocytic syndrome is a term used to describe proliferation of nonneoplastic (i.e., polyclonal), well-differentiated but highly erythrophagic macrophages. The condition is rare but has been recognized in dogs and cats. Unlike histiocytic sarcoma, which is neoplastic and in some forms also is characterized by highly erythrophagic macrophages, hemophagocytic syndrome is a secondary condition, occurring as a sequela of neoplasia, infection, or other underlying diseases. The macrophage proliferation and hyperactivation is a response to increased production of stimulatory cytokines occurring as part of the primary disease process. Macrophages are found in high numbers in the bone marrow and commonly in other tissues, including lymph nodes, spleen, and liver. Affected animals usually have cytopenias of two or more cell lines. Bone marrow findings reported in animals with hemophagocytic syndrome vary widely, ranging from hypoplasia to hyperplasia of cell lines with peripheral cytopenias, and may also include detection of macrophages containing phagocytosed hematopoietic precursor cells in addition to mature erythrocytes.
Hematopoietic Neoplasia
The term hematopoietic neoplasia encompasses a large, diverse group of clonal proliferative disorders of hematopoietic cells. This section is intended to help the student understand the basic biology (cell type, anatomic distribution, and biologic behavior) and diagnosis of hematopoietic neoplasia in veterinary medicine, with an emphasis on those forms that predominantly affect bone marrow, blood, and lymphoid tissue. Tumors of hematopoietic origin that predominantly affect other tissue, such as skin, are also mentioned briefly. Before listing specific diseases, it is worthwhile to consider the basic classification of hematopoietic neoplasia and mention some of the emerging diagnostic techniques used in veterinary medicine.
Hematopoietic neoplasia can be broadly classified as lymphoproliferative (lymphoid) or myeloproliferative (myeloid) disease, a distinction based on the fact that the earliest commitment of a pluripotent hematopoietic stem cell is to either a lymphoid or nonlymphoid lineage (see Fig. 13-3). Thus clonal proliferations of abnormal cells of lymphoid lineage are examples of lymphoid neoplasia, including the various forms of lymphoma (lymphosarcoma), lymphoid (lymphocytic) leukemia, and plasma cell tumors. Clonal proliferations of abnormal cells of nonlymphoid hematopoietic lineage (erythroid, granulocytic, monocytic and/or histiocytic, megakaryocytic), are considered myeloid neoplasms, including the various forms of myeloid (or myelogenous) leukemia, myelodysplastic syndrome, and histiocytic neoplasia. Mast cell tumors are not conventionally considered to be hematopoietic neoplasms, but in a technical sense they are also a form of myeloid neoplasia.
A number of techniques for diagnosing and classifying hematopoietic neoplasia are becoming increasingly available for routine use in veterinary medicine. For example, immunophenotyping refers to the use of antibodies recognizing specific molecules expressed on different cell types to determine the identity of a cell population of interest. Immunophenotyping on the basis of these lineage-specific or lineage-associated markers can be performed on histologic sections (immunohistochemistry), air-dried cytology smears (immunocytochemistry), or by laser analysis of cells in suspension in blood or buffer solutions (flow cytometry). Another emerging diagnostic technique is the clonality assay, a PCR test that can help identify neoplastic lymphoid proliferations on the basis of clonal rearrangements of genes encoding lymphocyte antigen receptors. In terms of practical application, the clonality assay is most useful in helping to distinguish lymphoid neoplasia from nonneoplastic lymphoid proliferations mimicking neoplasia.
In the last several decades, as new diagnostic techniques have become available, numerous systems for classifying hematopoietic neoplasia in humans have been proposed. These systems have been applied to veterinary medicine inconsistently but have helped to demonstrate the diversity of disease subtypes in animals. Some of these systems (the Rappaport and Kiel systems) did not attempt to correlate the classification of the neoplasm based on microscopic features with the clinical course of disease, whereas others (the Working Formulation for classification of lymphomas, sponsored by the National Cancer Institute) attempted to correlate survival data with morphologic criteria but did not take into account the cell of origin (i.e., B or T lymphocyte) of the tumor. A recently (2002) published system for classifying hematopoietic neoplasia in animals is an adaptation of the 2001 World Health Organization (WHO) system in humans. An effort is underway among a group of veterinary pathologists to coordinate the classification of hematopoietic neoplasia in humans and animals in a consistent, clinically relevant manner. The ultimate goal of this undertaking is to correlate pathologic findings (cell morphology, immunophenotype, tissue architecture, anatomic location) more precisely with clinical behavior of the neoplasm (prognosis, response to therapy).
The next section on hematopoietic neoplasia begins with leukemias and myelodysplastic syndromes, then covers the group of lymphoid malignancies known as lymphomas, and finishes by mentioning some other types of lymphoid and myeloid neoplasia.
Leukemia is an umbrella term referring to malignant hematopoietic neoplasms that originate in the bone marrow and typically have significant numbers of neoplastic cells in the blood. Hematopoietic neoplasms originating in the spleen, but primarily manifested in the blood, are also considered by some to be forms of primary leukemia. The term leukemia is also sometimes used to refer to other forms of neoplasia originating outside of the bone marrow or spleen, such as lymphoma, that progress to include significant bone marrow and blood involvement. For the purposes of this chapter, cases of secondary bone marrow or blood involvement are not considered leukemia but rather the “leukemic phase” of another primary neoplasm. Although primary leukemias usually have high numbers of neoplastic cells in circulation, this is not always the case. Some forms of leukemia (especially acute lymphoid leukemia in cats) have very low numbers of circulating neoplastic cells. This situation is sometimes known as aleukemic leukemia, although very close examination of blood smears usually reveals the underlying disease. The diagnosis may require confirmation by examination of the bone marrow, which in cases of leukemia contains disproportionately high numbers of neoplastic cells.
Leukemias are conventionally classified according to two criteria: as lymphocytic (lymphoid) or myelogenous (myeloid), based on the cell of origin, and as acute or chronic, based on the degree of differentiation of the neoplastic cells and their biologic behavior. This classification scheme is summarized in Table 13-3. Subcategories exist within each of these groups (e.g., lymphoid leukemias of B or T lymphocyte origin and myeloid leukemias of erythroid, granulocytic, monocytic, or megakaryocytic origin), as discussed further below.
TABLE 13-3
Basic Classification of Leukemias
| Lymphoid | Myeloid | |
| Chronic | CLL | CML |
| Acute | ALL | AML |
ALL, Acute lymphoid leukemia; AML, acute myeloid leukemia; CLL, chronic lymphoid leukemia; CML, chronic myeloid leukemia.
Chronic leukemias are proliferations of well-differentiated cells and are typically indolent, slowly progressive diseases. Often the number of neoplastic cells in circulation is very high. It is not unusual for the concentration to be in the hundreds of thousands per microliter. Mild to moderate nonregenerative anemia is common, but other cytopenias are not usually present. With chronic leukemias, the lineage of the cells is easy to identify by examining a blood or bone marrow smear. The cells look normal, or relatively normal, under the microscope. However, this can pose a diagnostic challenge because it is often not easy to tell whether the cells are a malignant population (i.e., leukemia) or a benign, reactive population (as in the case of marked inflammatory neutrophilia, reactive lymphocytosis, or secondary erythrocytosis), particularly when the concentration of cells in the blood is consistent with either neoplastic or nonneoplastic disease. Diagnosis of chronic leukemia in veterinary species is thus typically one of exclusion. An exception is when the concentration of cells in the blood is so high as to reasonably exclude the possibility of nonneoplastic disease (e.g., the concentration of lymphocytes in cases of chronic lymphocytic leukemia often exceeds 100,000 cells/µL). Necropsy findings depend on the stage of disease. In advanced cases with marked organ infiltration with neoplastic cells, there is often uniform splenomegaly, hepatomegaly, and lymphadenopathy. The bone marrow is highly cellular (Fig. 13-29) and histologically is densely cellular with well-differentiated neoplastic cells. In lymph nodes, there may be follicular lymphoid hyperplasia and neoplastic cells in the sinuses and medullary cords. Similarly the spleen may have follicular lymphoid hyperplasia and accumulation of neoplastic cells in the red pulp. The liver may have accumulations of neoplastic cells in the sinusoids and periportal areas.
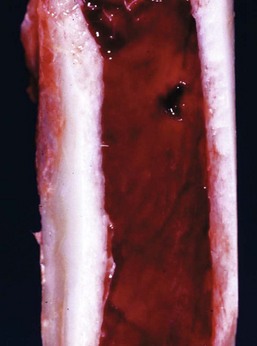
Fig. 13-29 Chronic leukemia, hypercellularity, bone marrow, dog.
Grossly, the marrow consists entirely of hematopoietic tissue (red) and no fat (yellow). In this case, hypercellularity of the marrow is attributable to neoplastic proliferation, but hyperplasia may cause similar gross findings. (Courtesy Dr. B.C. Ward, College of Veterinary Medicine, University of Mississippi and Noah’s Arkive, College of Veterinary Medicine, The University of Georgia.)
Chronic lymphoid leukemias (CLLs) are uncommon in veterinary species. The disease is rarely reported in horses and poorly characterized in cattle. In dogs, CLL is predominantly a disease of middle-aged to older dogs. Evidence indicates that most canine CLL cases are of T lymphocyte origin and that a subtype of granular T lymphocytes expressing CD8 (consistent with cytotoxic T lymphocyte lineage) is the most common form. Interestingly, most of these cases apparently originate in the spleen, with detectable bone marrow involvement occurring relatively late in the course of disease. In cats, evidence indicates that the majority of CLL cases have a T helper lymphocyte immunophenotype.
Chronic myeloid leukemias (CMLs) are rare in animals, with most reported cases occurring in dogs and cats. Types of CML reported in animals include chronic granulocytic leukemias of neutrophils, eosinophils, and basophils; chronic myelomonocytic leukemia; mast cell leukemia; essential thrombocythemia (CML of platelet lineage); and polycythemia vera (CML of erythrocyte lineage).
In contrast to chronic leukemias, acute leukemias are proliferations of poorly differentiated cells (Fig. 13-30) and typically have a rapid, aggressive clinical course. The distinction between chronic and acute leukemia is not always perfectly clear, either in terms of cell morphology or biologic behavior, and some cases fall “in the middle.” Moreover, chronic leukemia may develop into a more acute form (presumably because of additional mutation), a situation sometimes referred to as “blast crisis.” Acute leukemia has classically been defined by the presence of greater than 30% blast cells (large, immature cells) in the marrow or blood, although in human the threshold has been lowered to 20%. In general, the concentration of circulating neoplastic cells in acute leukemias does not tend to be as high as in chronic leukemias. Most animals with acute leukemia have cytopenias of two or more lineages, and pancytopenia (anemia, neutropenia, thrombocytopenia) is common. With acute leukemias, it is usually obvious from the presence of many large, immature cells in circulation that the patient has hematopoietic neoplasia. However, the cell of origin usually cannot be determined by routine microscopic examination because the cells are poorly differentiated. Identifying the specific cell type is best done by immunophenotyping and/or cytochemical staining. At necropsy, animals with acute leukemia have pale mucous membranes caused by anemia, and the bone marrow is highly cellular. Other findings depend on the degree to which neoplastic cells have infiltrated organs. There is often uniform splenomegaly, sometimes with infarcts. Histologically the bone marrow is densely cellular with poorly differentiated neoplastic cells. The splenic red pulp may be extensively infiltrated with neoplastic cells. The lymph nodes may have minor involvement of the medullary cords to diffuse involvement of the entire node.
Fig. 13-30 Acute leukemia, canine blood smear.
The specific lineage of the large, immature cells cannot be determined reliably based on routine staining. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Myelodysplastic syndrome (MDS) refers to a group of clonal myeloid proliferative disorders characterized by ineffective hematopoiesis. MDS does not meet the traditional definition of acute myeloid leukemia (AML) but is classically diagnosed based on these findings: less than 30% blasts in the bone marrow, cytopenias of more than one cell line, and morphologic evidence of dyshematopoiesis. Studies in people have shown that MDS also differs from AML on the molecular level (MDS is characterized by increased apoptosis compared with AML). MDS may lead to clinical illness and death as a result of the cytopenias and/or from transformation of the neoplasm into AML. MDS is rare in veterinary medicine, perhaps occurring most frequently in cats infected with FeLV.
The existing system for classification of AML and MDS in cats and dogs, an adaptation of the French-American-British (FAB) classification system in humans, was proposed by the Animal Leukemia Study Group (ALSG) in 1991 and generally has been accepted since then. However, the system does not include immunophenotyping or cytochemical staining characteristics, which have since become key aspects of classifying myeloid neoplasia in people.
The existing ALSG system in animals and the 2008 WHO classification system in humans are referenced in Web Box 13-2 and in the Suggested Readings list.
Acute lymphoid leukemia (ALL) is rarely recognized in horses and cattle. Cattle with lymphoid neoplasia caused by BLV infection often have high numbers of circulating neoplastic cells, but the disease is generally considered a form of lymphoma and is discussed next in the section on lymphoma. In a recent immunophenotypic study of 38 cases of ALL in dogs, 6 arose from B lymphocytes and 3 arose from CD8+ granular T lymphocyte. Cats with ALL often have low numbers of neoplastic cells in circulation, and examination of the bone marrow may be necessary to make the diagnosis.
AML occurs most often in dogs and cats but has also been reported in other domestic and wild animals. In dogs, the majority of acute leukemias are of myeloid or probable myeloid origin (21 of 38 cases in one recent study). MDS is recognized most often in cats, especially in animals infected with FeLV, but has also been reported in dogs and horses.
Lymphoma (Lymphosarcoma)
The term lymphoma (also known as lymphosarcoma or malignant lymphoma) encompasses a diverse group of malignancies arising in lymphoid tissue(s) outside the bone marrow. There are many different forms, with notable variability in the following:
• Anatomic location—multicentric, alimentary, mediastinal/thymic, other
• Immunophenotype—B lymphocyte versus T lymphocyte versus non-B/non-T lymphocyte
• Cellular morphology—size, nuclear features, mitotic rate
• Histologic pattern—diffuse versus follicular
• Biologic behavior—from low-grade (indolent) to high-grade (aggressive) tumors
As mentioned previously, there is an ongoing effort to develop an updated system for classifying hematopoietic neoplasia animals. This effort includes a specific project focusing on classification of lymphoma in dogs. The principle underlying this effort is that each category should constitute a distinct disease entity based on cell morphology, immunophenotype, tissue architecture, anatomic distribution, patient signalment, and full clinical history. A critical component of this effort is to test interobserver and intraobserver variability of interpretation.
Lymphoma is often suspected on the basis of organomegaly (e.g., generalized lymph node enlargement) or abnormal ultrasound findings (e.g., thickened intestines with abnormal echogenicity). Initial diagnosis typically is made based on aspiration cytology, biopsy and/or histopathology, or both, of the affected organ(s). In both cytologic and histologic preparations, lymphoma is characterized by a monomorphic population of morphologically atypical lymphocytes. Histologically, lymphoma is also characterized by disruption of normal tissue architecture. In general, lymphomas of small, well-differentiated cells with low mitotic rates are low-grade (indolent, slowly progressive) diseases, whereas lymphomas of large, poorly differentiated cells with high mitotic rates are high-grade (aggressive, rapidly progressive) diseases (Fig. 13-31). The type of tumor and extent of disease may be characterized further by immunophenotyping and clinical staging, respectively. Immunophenotyping is done to determine whether a tumor is of B lymphocyte or T lymphocyte origin (this distinction cannot be made reliably based on routine staining) (see Fig. 13-31). It is not necessarily the case that B lymphocyte lymphomas have a more favorable prognosis than T lymphocyte lymphomas because lymphomas of both cell types occur in low-grade and high-grade forms. Clinical staging includes examination of aspirates of bone marrow and other enlarged organs (e.g., spleen, liver, and other lymph nodes).
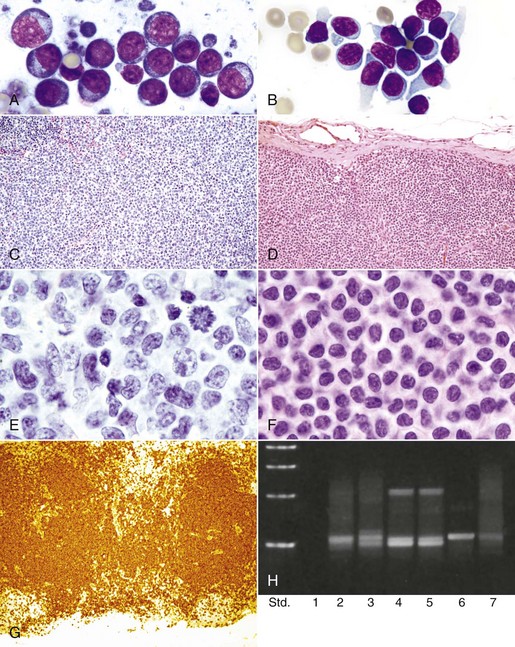
Fig. 13-31 Examples of high-grade and low-grade lymphoma (lymphosarcoma), lymph node, dog.
High-grade lymphoma: Fine-needle aspirate (A) and excisional biopsy (C and E) specimens. C and E are low- and high-magnification images, respectively, of the same specimen. Neoplastic lymphocytes are large and have large nuclei with dispersed chromatin and prominent nucleoli; a mitotic figure is evident at the top right of C. Low-grade lymphoma: Fine-needle aspirate (B) and excisional biopsy (D, F, and G) specimens, and results of polymerase chain reaction (PCR) testing of DNA extracted from the excisional biopsy. D and F are low- and high-magnification images, respectively, of the same specimen. In this sample, neoplastic lymphocytes are of small to intermediate size and have partially condensed chromatin; G shows a marked expansion of T lymphocytes. PCR results (H) confirm that these cells have a clonal T lymphocyte receptor gene rearrangement: Lane 1, negative control with no DNA; lanes 2 and 3, duplicates of a different case of T lymphocyte lymphoma; lanes 4 and 5, duplicates of this case; lane 6, positive control; lane 7, polyclonal control (i.e., normal). Wright’s stain (A and B), H&E stain (C-F), immunostain for T lymphocyte marker CD3 (G). (A courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee. B courtesy Dr. D.F. Edwards, College of Veterinary Medicine, University of Tennessee. C-G courtesy Dr. S.J. Newman, College of Veterinary Medicine, University of Tennessee. H courtesy Dr. W. Vernau, School of Veterinary Medicine, University of California, Davis.)
In cases of advanced disease, there may be marked bone marrow and blood involvement and distinguishing lymphoma from lymphoid leukemia can occasionally be difficult. The distinction can often be made on the basis of immunophenotyping, especially for CD34, which typically is expressed by neoplastic cells in acute leukemias and not in lymphomas. Other laboratory findings with lymphoma are variable. Mild-to-moderate nonregenerative anemia is probably the most common hematologic abnormality. Lymphopenia is noted more frequently than lymphocytosis, presumably because of abnormal trafficking of normal, nonneoplastic lymphocytes. Hypercalcemia is often associated with lymphoma caused by production of parathyroid hormone-related peptide (PTHrP) by the neoplastic cells. Such paraneoplastic hypercalcemia (humoral hypercalcemia of malignancy) may lead to pathologic calcification and related complications (e.g., renal failure).
Necropsy findings in cases of multicentric lymphoma include enlarged lymph nodes that bulge on cut section and are gray-white to light tan or reddish (see Fig. 13-80). In advanced cases, the normal architectural distinction between the cortex and medulla is obliterated. Infiltration of spleen and liver can result in diffuse enlargement or nodules. In alimentary lymphoma, affected portions of the GI tract are thickened and may be nodular. The lesions may be ulcerated. Enlarged mesenteric nodes may coalesce to form large masses. The gross appearance of thymic lymphoma is a large, gray, soft mass in the cranial mediastinum (see Fig. 13-49). Renal lymphoma often affects both kidneys and is evident as bilateral diffuse renomegaly. Central nervous system and bone marrow involvement of lymphoma can be difficult to detect on necropsy because of the similar gross appearance of neoplastic and normal tissue. In any of the various forms of lymphoma, large masses may have areas of central necrosis.
Lymphoma is the most common hematopoietic malignancy in animals and has been reported in all the domestic species.
Lymphoma is uncommon in horses, representing 5% or less of all neoplasms. Multicentric lymphoma is the most common form, with lesions occurring more frequently in the abdominal and thoracic cavities than in peripheral lymph nodes. Cutaneous and other forms may also occur. Lymph nodes in different anatomic regions (e.g., superficial, mediastinal, and alimentary) are often similarly affected, and infiltration of liver and spleen is common. Reports of the frequency of concurrent blood involvement are highly variable. The alimentary form of equine lymphoma is a wasting syndrome, presumably because of involvement of the small intestine, resulting in malabsorption. Cutaneous lymphoma in horses, as in other species, is a chronic disease without blood or internal organ involvement. A recent report of 37 cases of equine lymphoma found T lymphocyte lymphoma to be more common than B lymphocyte lymphoma. Weight loss and ventral edema were the most common clinical signs. Laboratory abnormalities present in at least one-third of cases included hyperfibrinogenemia, hypoalbuminemia, hyperglobulinemia, anemia, thrombocytopenia, and neoplastic cells in circulation. In all tumors, normal nodal architecture was diffusely effaced. Extensive necrosis was common. Many of the tumors were composed of morphologically heterogeneous cells, and a smaller subset had marked infiltration with histiocytes and multinucleated giant cells. Most T lymphocyte tumors were composed of cells of small to medium size, and most B lymphocyte tumors were classified as T lymphocyte-rich B lymphocyte neoplasms.
In cattle, lymphoma is primarily a multicentric B lymphocyte disease of adults and is strongly associated with BLV infection and BLV-related persistent lymphocytosis. Approximately 30% of cattle infected with BLV develop nonneoplastic persistent lymphocytosis and of these, less than 5% develop lymphoma. The BLV-induced form of lymphoma is known as bovine enzootic lymphoma. BLV is a retrovirus that persists within lymphocytes for the life of the infected animal. Transmission of the virus is mostly horizontal caused by transmission of infected lymphocytes as opposed to free virus in secretions. Blood-sucking arthropods or other mechanical means of transferring small numbers of infected lymphocytes (such as contaminated needles) are the principal means of spread. BLV infection is more common in dairy cattle than in beef cattle, presumably because dairy cattle husbandry favors viral transmission. Incidence of infection varies widely between regions and between herds. Antibodies against viral antigen can be detected by an agarose gel immunodiffusion test, and some herds remain free of BLV by testing and culling infected animals. In addition to lymph nodes, lymphoma in cattle often involves other locations, such as abomasum, vertebral canal, kidney, heart, retro-orbital space, and uterus (see Fig. 13-86). Enlargement of superficial lymph nodes is common, and enlarged pelvic and abdominal lymph nodes are often found on rectal palpation. Involvement of the digestive tract and heart may cause vagal indigestion or diarrhea and congestive heart failure, respectively. Lymphoma of the central nervous system is most frequently manifested as posterior paresis or paralysis because of pressure by the extradural mass on the cauda equina or on the lumbar cord. Other sporadic forms of lymphoma not associated with BLV infection also occur in cattle, including thymic, multicentric, and cutaneous forms affecting young animals. The thymic form of sporadic bovine lymphoma is characterized by large cranial thoracic and lower cervical masses, respiratory distress, and weight loss in cattle less than 2 years of age. The sporadic multicentric form is characterized by disseminated disease in calves 3 to 6 months of age. Affected calves frequently have widespread lymph node enlargement in addition to hepatic, splenic, and renal involvement. Marked blood and bone marrow involvement may also occur. The cutaneous form is rare, occurs in young cattle, and consists of discrete cutaneous plaques or large scabby lesions.
In dogs, lymphoma is a common form of neoplasia, representing approximately 7% to 9% of all malignancies and the large majority of hematopoietic malignancies. No retroviral or other cause is known. Lymphoma is typically a disease of middle-aged to older dogs. Multicentric lymphoma with generalized lymph node enlargement is the most common form (80% to 85% of all cases of canine lymphoma), and the majority of these (approximately 80%) are intermediate- or high-grade tumors. Other less common forms of canine lymphoma include alimentary, thymic, and cutaneous forms, and sporadically occurring forms in virtually every conceivable location. Approximately 70% to 80% of canine lymphomas are of B lymphocyte origin. Although the remaining cases are mostly of T lymphocyte origin, “null” (non-B, non-T, possibly of natural killer cell origin) lymphomas also occur. Clinical signs may be referable to the specific organ systems affected but are usually absent or nonspecific when lymphoma is first diagnosed. Approximately 15% of dogs with lymphoma and approximately 40% of those with mediastinal lymphoma, are hypercalcemic. In the majority of canine lymphoma cases, significant blood or bone marrow involvement is not detected by routine methods on initial diagnosis but may be seen with advanced disease. Splenic and hepatic involvement is common in cases of canine multicentric lymphoma.
In cats, lymphoma is among the many hematopoietic abnormalities caused by FeLV, and the epidemiology of feline lymphoma has changed with the development of routine vaccination and testing for FeLV. Cats infected with FeLV are likely to develop lymphoma at a younger age, and are more likely to develop certain types of lymphoma, especially mediastinal and multicentric T lymphocyte forms, than FeLV-negative cats. These predispositions still apply, but the percentage of cats infected with FeLV has decreased dramatically. As a result, mediastinal (thymic) and multicentric forms of lymphoma, which used to represent the majority of cases, have become relatively uncommon. At present, approximately 80% to 90% of cats with lymphoma are FeLV-negative, and the majority of lymphoma cases are the alimentary form. Other less common locations in cats include mediastinal, multicentric, renal, and other extranodal sites. FeLV-positive cats with lymphoma still tend to be younger, and their tumors more often of T lymphocyte origin, than FeLV-negative cats. The large majority (>80%) of cats with mediastinal lymphoma, and approximately one-third of cats with multicentric lymphoma, are FeLV-positive. Mediastinal and multicentric forms, although less common now, are still more likely to occur in younger cats, whereas the alimentary form typically develops in older cats (>10 years of age). The neoplastic cell type also tends to vary by anatomic location. Alimentary lymphoma in cats is predominantly a B lymphocyte disease, whereas mediastinal lymphoma is predominantly a T lymphocyte disease (consistent with thymic origin). A subtype of intestinal lymphoma in cats, large granular lymphocyte (LGL) lymphoma, is predominantly a T lymphocyte disease with a highly aggressive biologic behavior. Unlike dogs, cats are usually clinically ill when lymphoma is first diagnosed. In addition to nonspecific signs, such as weight loss, anorexia, and poor grooming habits, cats often have signs referable to the affected organ systems. For example, animals with alimentary lymphoma often have chronic diarrhea and vomiting and may have palpable abdominal masses, whereas those with mediastinal lymphoma are often dyspneic.
Lymphoma is the most commonly reported malignancy in pigs and may be more likely to affect females than males. Multicentric lymphoma is the most common form of the disease in pigs. Lymph node enlargement is more common in visceral than peripheral nodes. Other commonly affected organs include spleen, liver, kidney, and bone marrow. Lymphoma often affects animals less than 1 year of age, and the mediastinal form of lymphoma tends to affect younger pigs more commonly than the multicentric form. A viral cause (C type viruses) for lymphoma in pigs has been suggested, but transmission studies are lacking. A form of hereditary multicentric lymphoma has also been reported in inbred herds.
Plasma Cell Neoplasia
There are two main forms of plasma cells tumors recognized in veterinary species: multiple myeloma (MM) and plasmacytoma.
MM is a malignant tumor of plasma cells that arises in the bone marrow and usually secretes large amounts of Ig. Blood involvement of the neoplastic cells is not a feature of the disease. MM is a rare disease in animals. Dogs are affected more frequently than other species, but MM has also been reported in horses, cattle, cats, and pigs. The hallmark laboratory finding in patients with MM is hyperglobulinemia, which occurs because of production of large amounts of Ig or Ig subunit by the neoplastic cells. This homogeneous protein fraction is often called paraprotein or M protein. Concentrations of other Igs are often decreased.
Diagnosis of MM is based on finding a minimum of two or three (opinions vary) of the following abnormalities:
• Markedly increased numbers of plasma cells in the bone marrow, especially in aggregates (Fig. 13-32, A). A threshold value of plasma cells constituting at least 30% of nucleated cells in the marrow has been proposed as a diagnostic criterion. The neoplastic cells composing the tumor may be well-differentiated plasma cells or poorly differentiated cells with increased anisocytosis and anisokaryosis, visible nucleoli, and multinucleation.
Fig. 13-32 Multiple myeloma and monoclonal gammopathy.
A, Canine bone marrow aspirate. Many of the neoplastic plasma cells in the bone marrow aspirate have pink-tinged cytoplasm, the result of a high concentration of immunoglobulin. Wright’s stain. B, Multiple myeloma, cat. Agarose gels and densitometry tracings showing results of serum electrophoresis. The serum has a high concentration of a monoclonal immunoglobulin (the dark band [arrow] on the right of the gel, corresponding to the tall peak on the right of the tracing). C, Normal cat. Agarose gels and densitometry tracings showing results of serum electrophoresis. The serum has a normal distribution of protein fractions, the most abundant being albumin (the dark band [arrow] on the left of the gel, corresponding to the tall peak on the left of the tracing). (A courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee. B and C courtesy Dr. S.A. Kania, College of Veterinary Medicine, University of Tennessee.)
• Monoclonal gammopathy because of clonal production of Ig or Ig fragments by the neoplastic cells. Monoclonality is demonstrated by serum protein electrophoresis (Fig. 13-32, B) and can be characterized further using immunodiagnostic techniques (see later discussion). The term gammopathy is used because most Igs migrate in the γ-region of an electrophoresis gel, although some (especially IgA and IgM) may migrate in the β-region. Occasionally, biclonal or other atypical electrophoretic patterns may be seen with MM as a result of protein degradation, complex formation, or binding to other proteins, or when the tumor includes more than one clonal population. It is important to note that monoclonal gammopathy is not specific to MM but has also been reported in cases of B lymphocyte lymphoma and some nonneoplastic conditions such as canine ehrlichiosis or leishmaniasis. What appears to be a monoclonal pattern in the γ-region on routine serum electrophoresis may actually be oligoclonal (i.e., may include several Igs with a very similar migration pattern that are increased because of an immunologic response to antigenic stimulation). (This is unlikely to be the case with “spikes” in the β-region because nonneoplastic conditions typically do not result in high concentrations of IgA or IgM.) Definitively distinguishing monoclonal from oligoclonal patterns in the β-region requires immunoelectrophoresis or immunofixation using species-specific antibodies recognizing different Ig subclasses and subunits.
• Radiographic evidence of osteolysis. Work with human cell cultures has shown that osteoclasts support the growth of myeloma cells and that direct contact between the two cell types increases the myeloma cell proliferation and promotes osteoclast survival.
• Light chain proteinuria. Free Ig light chains (Bence Jones proteins) are of low molecular weight and pass through the glomerular filter into the urine. These proteins do not react with urine dipstick protein indicators and are most specifically detected by electrophoresis and immunoprecipitation.
Other pathologic findings in patients with MM may include the following:
• Hypercalcemia, most likely caused by increased osteoclast activity
• Lesions associated with marked hyperglobulinemia (hemorrhage caused by secondary platelet dysfunction, renal amyloidosis, and hyperviscosity syndrome)
• Cytopenias caused by high numbers of neoplastic cells in the bone marrow
MM typically has a slowly progressive clinical course. Common sites of metastasis include the spleen, liver, lymph nodes, and kidneys.
Cutaneous plasmacytomas are solid tumors of plasma cell origin that involve the skin or mucous membranes. Masses may be single or multiple. These tumors are usually benign, and excision is usually curative, but more aggressive forms may occur. Cutaneous plasmacytoma is discussed in more detail in Chapter 17. The tumor type known as extramedullary plasmacytoma (EMP) is a malignant solid tissue tumor of plasma cells arising from sites other than the bone marrow. The tumor is rare in animals, occurring most often in dogs and also reported in horses and cats. In one study, there was a disproportionately high percentage of EMP cases in cocker spaniels. The tumors occur most frequently in the GI tract but may also occur in the trachea, spleen, kidney, uterus, central nervous system, and elsewhere. Grossly, the tumors may be multinodular or cause thickening of the intestinal wall. Metastasis to regional lymph nodes is common. As with MM, the neoplastic cells composing the tumor may be well-differentiated to poorly differentiated (Fig. 13-33). EMPs produce monoclonal immunoglobulins, and monoclonal gammopathy has been reported in some cases of EMP. If there is bone or bone marrow involvement of a malignant plasma cell tumor, it is considered to be MM. Amyloidosis is associated with EMP in many reports and may be useful in distinguishing extramedullary plasmacytoma from other tumors. The distinction between cutaneous plasmacytoma and extramedullary plasmacytoma is not always clear, and benign plasmacytomas of the skin have also been referred to as cutaneous extramedullary plasmacytomas.
Histiocytic Neoplasia
Histiocytic sarcoma (HS) is an uncommon malignant neoplasm of histiocytic (macrophage or dendritic cell) origin. It occurs most frequently in the dog but has also been reported in the cat. There is a general belief that Rottweiler and Bernese mountain dogs are at increased risk for HS, although good epidemiologic data is thus far lacking. Some have called the disseminated form of this disease, with multiple organ involvement, by the name malignant histiocytosis, reserving the term histiocytic sarcoma for cases of single solid tumors, but there is an emerging consensus that the latter term encompasses both forms of the disease. The disseminated form of the disease has a rapid, aggressive clinical course. Sites commonly involved include the spleen, lung, lymph nodes, bone marrow, skin, and subcutis. Liver involvement occurs secondary to disease in the spleen. The solitary form of HS can occur at any of the aforementioned sites and also in joints (in close association with the subsynovium) or brain. Most cases of HS are of dendritic antigen-presenting cell origin, with a similar immunophenotype to (but dramatically different biologic behavior than) cutaneous histiocytoma. Fewer cases of HS are of macrophage cell origin. These malignant cells have an immunophenotype characteristic of resident macrophages in the splenic red pulp and the bone marrow, and frequently show marked phagocytosis of erythrocytes (hemophagocytosis). Dogs with this form of HS frequently have a hemophagocytic syndrome characterized by severe nonregenerative to mildly regenerative anemia (presumably a result largely of destruction of RBCs by the neoplastic cells), splenomegaly, hepatomegaly, and extramedullary hematopoiesis in the spleen and elsewhere. Microscopically, HS tumor cells are large, round-to-spindloid in shape and vary from relatively well-differentiated histiocytic morphology to cells with notable features of malignancy (Fig. 13-34).
Fig. 13-34 Histiocytic sarcoma, canine mesenteric lymph node aspirate.
The neoplastic cells are round cells with abundant gray-blue cytoplasm. Note the following features of malignancy: A, The low-magnification image shows notable variation in cell and nuclear sizes. B, The high-magnification image shows multinucleation, prominent nucleoli, and mitotic figures, as well as a background population of lymphocytes. Wright’s stain. (Courtesy Dr. M.M. Fry, College of Veterinary Medicine, University of Tennessee.)
Cutaneous histiocytosis (CH) and systemic histiocytosis (SH), nonneoplastic canine immunoregulatory disorders that share some clinical features of histiocytic malignancies, are covered in the section on canine immune-mediated diseases, and CH is discussed in more detail in Chapter 17.
Canine cutaneous histiocytoma, a benign neoplasm of epidermal Langerhans cell origin, and feline progressive histiocytosis, an initially indolent cutaneous neoplasm that is probably of dendritic cell origin, are discussed in detail in Chapter 17.
Mast Cell Neoplasia
Mast cell tumors (MCTs) of the skin and other organs are relatively common in animals, especially in dogs, and are covered in Chapter 6. Primary leukemia of mast cell origin is a rare form of myeloid leukemia. Mast cells normally are not present in circulation, but the finding of mast cells in the blood (mastocytemia) does not necessarily indicate myeloid neoplasia. In fact, one study found that severity of mastocytemia in dogs was frequently higher in animals without MCTs than those with MCTs and that random detection of mast cells in blood smears usually is not a result of underlying MCT.
Hemostasis
This section covers disorders of other components of hemostasis such as coagulation factors. Information on this topic is available at evolve.elsevier.com/Zachary/McGavin/. Platelet disorders are covered in the sections on disorders of bone marrow and blood cells.
Congenital Disorders
Hemophilia A refers to an inherited deficiency of functional coagulation factor VIII and has been recognized in horses, cattle, dogs, and cats. Hemophilia B, also known as Christmas disease, refers to an inherited deficiency of functional coagulation factor IX and has been recognized in dogs and cats. Both forms of hemophilia have an X-linked recessive mode of inheritance. Affected males have variable tendencies to bleed, depending on the severity of the deficiency, exposure to trauma, and size and activity level of the affected individual. Carrier females are usually asymptomatic.
Other Inherited Factor Deficiencies
Factor XI deficiency is the most common inherited coagulopathy in cattle and has also been reported in dogs. The mode of inheritance is probably autosomal recessive. Reports describe excessive bleeding in affected individuals after surgery. The most commonly affected dog breed is the Kerry blue terrier. In these dogs, hemorrhage is often delayed, sometimes developing several days after surgery. Other factor deficiencies also have been reported in animals. These conditions include deficiencies of factor VII (reported in dogs), factor XII (relatively common in cats, also reported in dogs), and prekallikrein (reported in dogs and horses). In most cases, affected individuals are asymptomatic or have only mild clinical bleeding tendencies but may have excessive bleeding after trauma or surgery.
Other Inherited Coagulopathies
A defect in γ-glutamyl carboxylase, the enzyme required for normal carboxylation of vitamin K–dependent clotting factors, has been recognized as the cause of an inherited coagulopathy in Rambouillet sheep.
von Willebrand disease
von Willebrand disease (vWD) actually refers to a group of inherited conditions characterized by a quantitative or qualitative deficiency of vWF, which is a soluble multimeric glycoprotein that circulates as a complex with and stabilizes coagulation factor VIII and mediates binding of platelets to subendothelial collagen (see Fig. 2-18). Although not technically a platelet disorder, vWD is often classified as such because it results in a loss of normal platelet function. vWD is the most common canine hereditary bleeding disorder and has also been described in many other domestic species. Different types of vWD vary in terms of mode of inheritance and severity of clinical disease. Type I vWD is characterized by low plasma vWF concentration but normal multimeric proportions and a mild-to-moderate clinical bleeding tendency; it has been reported in many dog breeds. Type II vWD is characterized by low vWF concentration, absence of large multimers, and a moderate-to-severe bleeding tendency; it has been reported in German shorthaired pointer and German wirehaired pointer dogs. Type III vWD is characterized by absence of vWF and a severe bleeding tendency; familial and sporadic cases have been reported in numerous dog breeds.
Toxicoses
Antagonism of vitamin K leads to production of a nonfunctional form of some coagulation factors, and resulting coagulopathy; a similar condition results from vitamin K deficiency. A number of coagulation factors—factors II, VII, IX, and X (collectively known as the vitamin K-dependent factors), as well as the regulatory molecules Protein C and Protein S—must undergo carboxylation to be functional. This posttranslational modification is catalyzed by the enzyme γ-glutamyl carboxylase and requires vitamin K as a cofactor. Vitamin K is oxidized during the carboxylation reaction and is converted back into its active reduced form by the enzyme vitamin K epoxide reductase. Conditions associated with avitaminosis K include poisoning with coumarin-related molecules; fat malabsorption (vitamin K is one of the fat-soluble vitamins) caused by primary intestinal disease or impaired biliary outflow (uncommon); dietary deficiency (rare); and antibiotics that interfere with vitamin K absorption or usage. Coumarin-related rodenticides, such as warfarin, act by inhibiting vitamin K epoxide reductase, resulting in an absence of vitamin K in its active reduced form (Web Fig. 13-2). This inhibition lasts until the rodenticide is metabolized and cleared. How long this takes depends on the half-life ( ) of the rodenticide and dose, but it may take many weeks. Second-generation rodenticides, such as bromadoline and brodifacoum, are more potent than warfarin, with a longer
) of the rodenticide and dose, but it may take many weeks. Second-generation rodenticides, such as bromadoline and brodifacoum, are more potent than warfarin, with a longer  . Mycotoxins in the same family as anticoagulant rodenticides are associated with mildly sweet clover or sweet vernal grass and cause coagulopathy by the same mechanism. Laboratory findings include prolonged coagulation times (prothrombin time [PT], partial thromboplastin time [PTT], and activated clotting time [ACT]). Early in the course of rodenticide and related toxicoses, PT may be the only one of these tests that is prolonged, because factor VII has the shortest
. Mycotoxins in the same family as anticoagulant rodenticides are associated with mildly sweet clover or sweet vernal grass and cause coagulopathy by the same mechanism. Laboratory findings include prolonged coagulation times (prothrombin time [PT], partial thromboplastin time [PTT], and activated clotting time [ACT]). Early in the course of rodenticide and related toxicoses, PT may be the only one of these tests that is prolonged, because factor VII has the shortest  of the vitamin K–dependent factors, but the other tests become prolonged as nonfunctional forms of the other factors accumulate. In uncomplicated cases, patients are not thrombocytopenic. A wide range of hemorrhagic lesions may occur in affected individuals, including ecchymoses, epistaxis, gingival bleeding, hematomas, hemoptysis, melena or hematochezia, hematuria, and other forms of hemorrhage. The treatment for cases of rodenticide and related toxicoses is regular administration of exogenous vitamin K1 until the toxin is cleared (determined by repeat coagulation testing after withholding treatment).
of the vitamin K–dependent factors, but the other tests become prolonged as nonfunctional forms of the other factors accumulate. In uncomplicated cases, patients are not thrombocytopenic. A wide range of hemorrhagic lesions may occur in affected individuals, including ecchymoses, epistaxis, gingival bleeding, hematomas, hemoptysis, melena or hematochezia, hematuria, and other forms of hemorrhage. The treatment for cases of rodenticide and related toxicoses is regular administration of exogenous vitamin K1 until the toxin is cleared (determined by repeat coagulation testing after withholding treatment).
Disseminated Intravascular Coagulation (DIC)
DIC is a syndrome characterized by continuous activation of both coagulation and fibrinolytic pathways and also known as consumptive coagulopathy. DIC is not a primary disease process but rather a secondary complication of many types of underlying disease. It is common in critically ill domestic animals. DIC involves an initial hypercoagulable phase, resulting in thrombosis and ischemic tissue damage, and a subsequent hypocoagulable phase as a result of consumption of coagulation factors and platelets, resulting in hemorrhage (Web Fig. 13-3). The list of underlying conditions associated with DIC includes neoplasia, sepsis, endotoxemia, immune-mediated disease, intravascular hemolysis, shock, heat stroke, and obstetric complications. The pathogenesis of DIC typically involves the release of tissue factor (thromboplastin) and subsequent activation of coagulation pathways and platelets but may also involve defective normal inhibition of coagulation or defective fibrinolysis. Classically, diagnosis of DIC is based on clinical evidence of hemorrhage and/or thromboembolic disease and a triad of laboratory findings: thrombocytopenia, usually moderate (below the lower reference value but above 50,000/µL); prolonged coagulation times (PT and/or PTT); and decreased fibrinogen, or increased concentration of plasma fibrin degradation products or D-dimer. Milder forms of DIC that do not meet all of the diagnostic criteria also occur. Decreased plasma antithrombin (antithrombin III) concentration and schistocytosis are other laboratory abnormalities often found in patients with DIC.
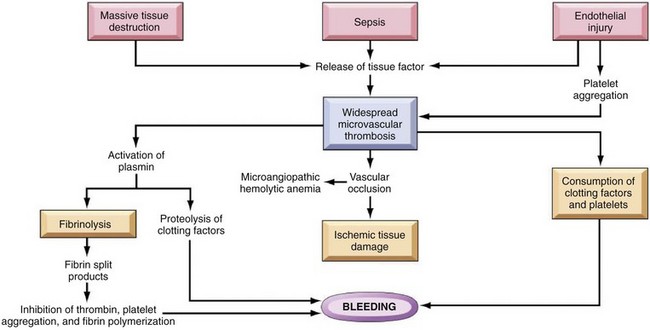
Disorders of Horses
Flavin Adenine Dinucleotide Deficiency
Flavin adenine dinucleotide (FAD) is a cofactor for cytochrome b5 reductase, the enzyme that maintains hemoglobin in its functional reduced state, and for the glutathione reductase reaction that also helps protect erythrocytes from oxidative damage. Deficiency of erythrocyte FAD caused by abnormal erythrocyte riboflavin metabolism was recently reported in a Spanish mustang mare with methemoglobinemia and eccentrocytosis and has also been recognized in a Kentucky mountain saddle horse gelding.
Infectious Diseases
Equine Infectious Anemia Virus
EIAV is a lentivirus that infects cells of the mononuclear phagocyte system in horses (also ponies, donkeys, and mules) throughout the world. Natural transmission is by arthropods, and the virus can also be transmitted transplacentally. Disease occurs in both an acute, potentially fatal form after initial infection, as well as a chronic intermittent form associated with recurrent viremia. Recurrent episodes happen mostly in the first year after infection, and tend to decrease in frequency and severity with time. Infection is lifelong and animals may become asymptomatic carriers. Viral antigen is found mainly within liver, spleen, and serum of infected animals, and also in bone marrow, lymph nodes, thymus, circulating mononuclear cells, and other tissues. EIAV causes anemia by both immune-mediated hemolysis and decreased erythropoiesis. Hemolysis is typically extravascular but may have an intravascular component during the acute phase. Decreased erythropoiesis may result from direct suppression of early-stage erythroid cells by the virus, as well as anemia of inflammation. Thrombocytopenia, likely caused by secondary immune-mediated destruction, is also a classic feature of acute EIAV infection and recurring febrile episodes. Clinical findings associated with viremic episodes include fever, depression, icterus, petechial hemorrhages, lymph node enlargement, and dependent edema. EIA infection is diagnosed on the basis of the Coggin’s test, an agarose gel immunodiffusion test for the presence of the antibody against the virus.
Animals dying during hemolytic crises have icterus, anemia, and widespread hemorrhages. The spleen and liver are enlarged, dark and turgid, and they and other organs have superficial subcapsular hemorrhages. Petechiae are evident beneath the renal capsule and throughout the cortex and medulla. The bone marrow is dark red as a result of replacement of fat by hematopoietic tissue; the extent of replacement is an indication of the duration of the anemia. The severity of microscopic lesions is dependent on the chronicity of the disease, and they are most significant in the spleen, liver, and bone marrow. As would be anticipated, microscopic findings of the spleen are predominantly influenced by the number and activity of macrophages, which is a reflection of the duration of the disease and the frequency of hemolytic episodes. Hemosiderin-laden macrophages persist for months to years; therefore, large numbers are consistent with chronicity. Kupffer cell hyperplasia with hemosiderin stores and periportal infiltrates of lymphocytes are the most significant changes in the liver. Bone marrow histologic findings vary depending on the duration of the disease. In most animals, the marrow is cellular because of the replacement of fat by intense, orderly erythropoiesis. Granulocytes are relatively less numerous and plasma cells are increased. As in the spleen, hemosiderin-laden macrophages are present in large numbers in chronic cases. In more chronic cases, emaciated animals have serous atrophy of fat (see Web Fig. 13-1).
Disorders of Ruminants (Cattle, Sheep, and Goats)
Congenital Dyserythropoiesis in Polled Herefords
A syndrome of congenital dyserythropoiesis and alopecia occurs in polled Hereford calves. The disease is often fatal. Hematologically, the condition is characterized by anemia that is nonregenerative to mildly regenerative (poorly regenerative given the degree of anemia). Bone marrow findings include marked erythroid predominance and morphologic abnormalities, consistent with ineffective erythropoiesis. The specific defect has not been identified.
Nutritional and Metabolic Disorders
Hypophosphatemic Hemolytic Anemia
Marked hypophosphatemia is recognized as a cause of intravascular hemolytic anemia in postparturient dairy cows. Hypophosphatemia develops in these animals because of increased loss of phosphorus in their milk. Biochemical studies suggest that the mechanism of hemolysis involves decreased erythrocyte production of ATP, which may lead to compromised membrane and cytoskeletal integrity. An accompanying decrease in reducing capacity and increase in methemoglobin concentration have also been noted in experimental studies of hypophosphatemic hemolytic anemia in dairy cattle, suggesting that oxidative mechanisms may also contribute to anemia. Affected cows are anemic and hemoglobinuric. Gross postmortem findings include pallor, decreased viscosity of the blood, and discolored pale yellow and swollen liver and kidney. Renal tubular necrosis and hemoglobin pigment within the tubules is evident microscopically. Hemolysis has also been reported in association with hypophosphatemia in dogs and cats.
Infectious Diseases
Bovine Viral Diarrhea Virus
Thrombocytopenia, often severe, has been reported in association with acute bovine viral diarrhea virus (BVDV) infection in calves and adult cattle. Infection with type II BVDV has been specifically associated with a thrombocytopenic hemorrhagic syndrome. Calves infected with type II BVDV have also been shown to have impaired platelet function. Investigations of the mechanism of BVDV-induced thrombocytopenia have resulted in varying, sometimes conflicting, conclusions. More than one study has shown viral antigen associated both with bone marrow megakaryocytes and with circulating platelets. Evidence of impaired thrombopoiesis (megakaryocyte necrosis, megakaryocyte pyknosis, and degeneration) and increased thrombopoiesis (megakaryocytic hyperplasia, increased numbers of immature megakaryocytes) in the bone marrow has been reported in type II BVDV-infected animals, including concurrent megakaryocyte necrosis and hyperplasia in some experimental subjects. To our knowledge, antibody-mediated destruction of platelets has not been shown.
Disorders of Dogs
Cyclic Hematopoiesis
Cyclic hematopoiesis is an inherited disorder of pluripotent hematopoietic stem cells, recognized in dogs and people. In dogs, the condition has an autosomal recessive inheritance pattern and is associated with dilute hair coat color. In dogs, cyclic hematopoiesis (also known as lethal gray collie disease) is characterized by predictable fluctuations in concentrations of blood cells that occur in 14-day cycles. The pattern is cyclic marked neutropenia and in a different phase, cyclic reticulocytosis, monocytosis, and thrombocytosis. Production of key cytokines involved in regulation of hematopoiesis is also cyclic. The specific lesion in cyclic hematopoiesis is believed to involve defective intracellular signaling but remains to be explained on the molecular level. Neutropenia predisposes affected animals to infection, and many die of infectious causes. Other related clinical manifestations include bleeding tendency, attributable at least in part to defective platelet function, and systemic amyloidosis, which occurs because of cyclic increases in concentration of acute phase proteins during phases of monocytosis.
Phosphofructokinase Deficiency
Inherited autosomal recessive deficiency of the erythrocyte glycolytic enzyme, phosphofructokinase (PFK), is described in English springer spaniel, American cocker spaniel, and mixed-breed dogs. The enzyme is also deficient in muscle tissue of affected dogs. Erythrocytes in PFK-deficient dogs have decreased ATP and 2,3-diphosphoglycerate production, and increased fragility under alkaline conditions. The disease is characterized by chronic, extravascular hemolysis with marked reticulocytosis. The marked regenerative response may compensate for the ongoing hemolysis, therefore affected animals are not necessarily anemic. However, acute intravascular hemolytic episodes may occur with hyperventilation and resulting alkalemia. There are three genes encoding PFK enzymes, designated M in muscle and erythrocytes, L in liver, and P in platelets. A point mutation in the gene coding for the M enzyme results in an unstable, truncated molecule. A single DNA-based test is available to detect the common mutation.
Erythrocyte Structural Abnormalities
A heterogeneous group of conditions, known as hereditary stomatocytosis because of the characteristic slit-shaped area of central pallor in RBCs on stained blood smears, has been described in several dog breeds. The clinical manifestations vary in affected animals, and the specific underlying defects have not been identified. In all cases, however, erythrocytes have increased osmotic fragility and decreased survival.
Other (presumably heritable) erythrocyte abnormalities in dogs that are not associated with clinical signs include elliptocytosis caused by band 4.1 deficiency or β-spectrin mutation, and familial macrocytosis and dyshematopoiesis in Poodles.
Scott-Like Syndrome
An inherited thrombopathy resembling Scott syndrome in humans, in which platelets lack normal procoagulant activity, has been recognized in a family of German shepherd dogs. Affected dogs have a mild to moderate clinical bleeding tendency characterized by epistaxis, hyphema, intramuscular hematoma formation, and increased hemorrhage associated with surgery. The specific defect in these dogs has not been identified on the molecular level, but involves impaired expression of phosphatidylserine on the platelet surface.
Breed-Related Platelet Abnormalities
Cavalier King Charles spaniels often have a lower than normal concentration of platelets, many of which are abnormally large (a condition known as macrothrombocytopenia). The molecular basis for this trait is a mutation in the gene encoding β1 tubulin. In general, affected dogs are asymptomatic. Cavalier King Charles spaniels have been shown to have abnormal platelet aggregation in vitro in some studies, although the clinical significance of these findings, including a possible causal relationship between platelet abnormalities and mitral valve disease in this breed, is not clear.
Greyhounds tend to have lower concentrations of circulating platelets than other dog breeds, although this is not of any known clinical significance. Other Greyhound traits include lower neutrophil and plasma protein concentrations and higher hematocrits than other dog breeds.
Immune-Mediated Disorders
Cutaneous and Systemic Histiocytoses
CH, a rare immunoregulatory disorder caused by proliferation of activated dermal dendritic cells, is discussed in more detail in Chapter 17. SH is a familial disorder of Bernese mountain dogs (also found sporadically in other breeds) characterized by large, dense proliferations of activated interstitial dendritic cells in multiple tissues, including skin, peripheral lymph nodes, and ocular and nasal mucosa. The proliferating cells are immunophenotypically identical to those in CH, and the angiocentric skin lesions in SH are histologically identical to those in CH. Lesions of SH in locations other than skin are also angiocentric infiltrates consisting mostly of histiocytes and lymphocytes.
Disorders of Cats
Increased Erythrocyte Osmotic Fragility
A condition characterized by increased erythrocyte osmotic fragility has been described in Abyssinian and Somali cats. The specific defect has not been identified, but pyruvate kinase deficiency (which has been reported in these breeds) was excluded as the cause. Affected cats have chronic intermittent severe hemolytic anemia and often have other lesions secondary to hemolytic anemia (e.g., splenomegaly and hyperbilirubinemia).
Infectious Diseases
Cytauxzoon felis is a protozoal organism causing severe, often fatal, disease in cats. Cytauxzoonosis in domestic cats is relatively common in the South Central United States, particularly during summer months. Bobcats (Lynx rufus), and perhaps other wild felids, are believed to be the reservoir host. They are usually asymptomatic, with persistent parasitemia and transient schizogony. Cytauxzoon felis is transmitted by a tick vector, Dermacentor variabilis, which is probably essential for infectivity of the organism. Cytauxzoonosis has a schizogenous phase involving macrophages throughout the body (especially liver, spleen, lung, lymph nodes, and bone marrow) that causes systemic illness, and an erythrocytic phase that causes anemia of variable severity. Schizont-containing macrophages (Fig. 13-35) enlarge and accumulate within the walls of veins, eventually causing occlusion of the vessels. Merozoites are released and enter erythrocytes. Parasitemia develops relatively late in the course of infection. The signet ring–shaped erythrocytic inclusions (piroplasmas) of Cytauxzoon felis closely resemble small form Babesia (see Fig. 13-23, A) and some Theileria organisms. Affected cats typically become acutely ill with fever, pallor, and icterus and usually die within 2 to 3 days. For many years, cytauxzoonosis was considered to be almost always fatal. However, a recent report, in which numerous cats from a subregion of the endemic area in the United States survived infection with an organism with greater than 99% homology to Cytauxzoon felis, suggests the emergence of a less virulent strain. In affected cats, erythrophagocytosis is also often a prominent finding in tissue with the schizogenous phase.
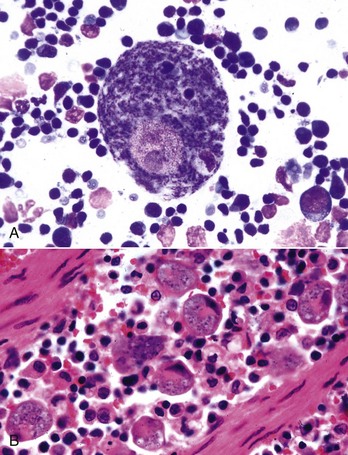
Fig. 13-35 Cytauxzoonosis, tissue aspirates, cat.
A, Lymph node aspirate. A large macrophage (center of figure) is laden with schizonts of Cytauxzoon felis. Wright’s stain. B, Splenic macrophages are filled with Cytauxzoon organisms. H&E stain. (A courtesy Dr. D.F. Edwards, College of Veterinary Medicine, University of Tennessee. B courtesy Dr. A.R. Doster, University of Nebraska; and Noah’s Arkive, College of Veterinary Medicine, The University of Georgia.)
Feline Leukemia Virus
FeLV is an oncogenic, immunosuppressive lentivirus associated with hematologic abnormalities of widely varying type and severity, including anemia in most infected cats. Manifestations of disease caused by FeLV infection vary depending on dose, viral genetics, and host factors, but normal hematopoiesis is probably suppressed to some degree in all cases. The virus infects hematopoietic precursor cells soon after the animal is exposed and continues to replicate in hematopoietic and lymphatic tissue of animals that remain persistently viremic. Persistently viremic cats are immunosuppressed and are prone to developing other diseases, including infectious diseases, bone marrow disorders, and lymphoma (lymphosarcoma).
FeLV-induced anemia is usually nonregenerative, presumably caused by direct effects of the virus on infected erythroid cells. However, macrocytosis and metarubricytosis (the presence of nucleated RBC precursors in circulation) are often noted in the absence of significant reticulocytosis—findings consistent with dyserythropoiesis, although the exact mechanism is not clear. Bone marrow of cats with FeLV-induced anemia often has evidence of arrested or disordered maturation of hematopoietic precursors. The relatively uncommon subgroup C viruses are associated with erythroid aplasia, probably because of infection of early stage erythroid precursors. Regenerative anemia may also occur with FeLV infection, often because of coinfection with Mycoplasma haemofelis.
FeLV infects hematopoietic cells and can produce a wide array of hematologic and other disease manifestations. FeLV may be detected in megakaryocytes and platelets in infected cats, and may result in platelet abnormalities, including thrombocytopenia, thrombocytosis, increased platelet size, and decreased function. Proposed mechanisms of FeLV-induced thrombocytopenia include direct cytopathic effects, myelophthisis, and immune-mediated destruction. Platelet life span and function have been shown to be decreased in FeLV-positive cats.
Feline Immunodeficiency Virus
FIV, another feline lentivirus, is associated with anemia in a minority of infected cats. Immunosuppressive effects of FIV from thymic depletion are discussed elsewhere. It is generally accepted that anemia does not result directly from FIV infection but instead develops because of concurrent disease such as co-infection with FeLV or hemotropic mycoplasma, other infection, or malignancy. The severity and type of anemia in FIV-infected cats depends on the other specific disease processes involved.
Lymphatic System
The philosophy of this book is to describe the response of an organ or tissue to injury, based on its anatomic components and cell types. This approach works well with most organs and facilitates the reader’s understanding of the pathologic processes. However, in the case of the lymphatic system, in which both the anatomic structures and immunologic functions are complex and interrelated, it is more difficult. Chapter 5 on the diseases of immunity describes some of these processes and to reduce repetition, the reader is referred to that chapter where appropriate. To understand the sequence of events in both normal and pathologic secondary lymphoid organs, it is desirable that the reader have an understanding of the three-dimensional anatomy of the spleen and lymph node, and particularly the pathways of the flow of blood, lymph, and lymphocytes through the lymph node and blood and lymphocytes through the spleen.
Under the classification of the Nomina Anatomica Veterinaria, the thymus, spleen, lymph nodes, and lymph nodules are part of the lymphatic system. They are also part of the immune system. Primary lymphoid organs are defined as the sites at which the component cells of the immune system (B and T lymphocytes) are formed. In most animals, there are two primary lymphoid organs (thymus and bone marrow), but in ruminants and birds, Peyer’s patches (ileum) and the bursa of Fabricius, respectively, are sites of B lymphocyte production. The spleen, lymph nodes, and lymph nodules are secondary lymphoid organs and are responsible for the immune responses (i.e., production of antibody and cell-mediated immunity). Thus ruminant Peyer’s patches are both primary and secondary lymphoid organs. However, because the spleen and lymph nodes contain numerous cells of the monocyte-macrophage system, they also have functions outside those of the immune system such as phagocytosis of particulate material (e.g., carbon, bacteria, and in the case of the spleen, senescent and altered erythrocytes).
The bone marrow is described in the first section of this chapter. The remaining primary lymphoid organ, the thymus, is described first in this section, followed by the secondary lymphoid organs: spleen, lymph nodes, and diffuse and nodular lymphatic tissues.
Thymus
The thymus is essential for the development and function of the immune system, specifically for the differentiation, selection, and maturation of T lymphocytes. This is described in detail in Chapter 5, and the interrelationship of the T lymphocytes with the components of the thymus are described in this section. The shape and location of the thymus vary among young domestic animals. In ruminants and pigs, the thymus has two lobes: cervical and thoracic. The cervical lobe is large and extends along the lateral surfaces of the cervical trachea. The size of the cervical lobe varies in cats and horses but is usually small. Dogs do not have a cervical lobe. The thoracic lobe is present in all domestic animals and lies in the cranial mediastinum, ventrally in the horse, pig, and dog and dorsally in ruminants.
The thymus has also been called a lymphoepithelial organ because besides lymphocytes it has an epithelial component. Histologically, it is divided into two portions, the stromal portion and the thymocyte portion (T lymphocytes at different stages of maturation and differentiation). All T lymphocytes in the thymus are derived from progenitor T lymphocytes in the bone marrow. The capsule, trabeculae, and blood vessels arise from the mesoderm of a branchial arch. Some stroma is designated “epithelial reticulum,” reflecting its embryonic origin from branchial pouches and the third pharyngeal cleft. It produces no reticular fibers and thus is different from the reticulum composed of a meshwork of specialized fibroblast-like cells with long interdigitating filamentous projections (reticular cells), through which are dispersed macrophages and occasional myoid cells (smooth muscle-like).
The lymphoid component is composed of differentiating lymphocytes, which during gestation arise from hematopoietic progenitor cells, initially in the fetal liver, then spleen and later in the fetal bone marrow. The epithelial component consists of individual cells in the cortex and medulla and aggregates of epithelial cells in the medulla. The latter, called Hassall’s corpuscles, are a characteristic histologic feature of the thymus. Cortical and medullary epithelial cells contribute to the microenvironment and are essential for differentiation, selection, and maturation of T lymphocytes in neonate and adult thymuses.
Microscopically, the thymus is divided into incomplete lobules, each of which has a cortex and a medulla, with the medullas being confluent centrally (Fig. 13-36). On the basis of functional and epithelial components, three zones are recognized: a subcapsular zone, a cortex, and a medulla. Bone marrow–derived T lymphocytes enter the circulation, travel to the thymus, and enter at the subcapsular zone (Fig. 13-37). Here, they begin differentiation and selection processes and develop into mature naïve T lymphocytes as they traverse the thymic cortex to the medulla. In the cortex, T lymphocytes that recognize self-molecules (i.e., major histocompatibility complex [MHC] molecules) but not self-antigens are permitted to mature by a process called positive selection. Cells that do not recognize MHC molecules are removed by apoptosis. Those T lymphocytes that recognize both MHC molecules and self-antigens are removed by macrophages at the corticomedullary junction, a process called negative selection. Because of the rigid differentiation requirements attributable to MHC restriction and tolerance (positive and negative selection, respectively) only a small fraction (<5%) of the developing T lymphocytes that arrive at the thymus from the bone marrow survive. Mature naïve T lymphocytes exit the thymus through postcapillary venules in the corticomedullary region, enter the circulation, and recirculate through secondary lymphoid tissues in a highly regulated manner. The T lymphocyte regions of secondary lymphoid organs are specialized sites where these circulating mature naïve T lymphocytes (T lymphocytes that have not been exposed to their T lymphocyte receptor-specific antigen) are activated. On exposure to their specific antigens, they undergo additional phases of development and differentiate into effector and memory cells, some of which remain localized in lymph nodes and others migrate to sites of infection. The balance between the rates of production and distribution of T lymphocytes is critically important in maintaining immune homeostasis. In fact, immunodeficiency or autoimmunity may result from inadequate or overly zealous activity, respectively, of a particular subset of T lymphocytes.
Fig. 13-36 Schematic illustration of the organization of the thymus.
The thymus consists of several incomplete lobules. Each lobule contains an independent outer cortical region, but the central medullary region is shared by adjacent lobules. Trabeculae, extensions of the capsule down to the corticomedullary region, form the boundary of each lobule. The cortex consists of stromal cells and developing T lymphocytes (thymocytes), macrophages, and cortical epithelial cells. Major histocompatibility complex class I and II molecules are present on the surface of the cortical epithelial cells. The characteristic deep blue nucleus staining of the cortex in histological preparation reflects the predominant population of T lymphocytes as compared with the less basophilic medulla, which contains a lower number of thymocytes. Hassall’s corpuscles are a characteristic component of the medulla and are not seen in the cortex. (From Kierszenbaum AL: Histology and cell biology: an introduction to pathology, ed 2, St Louis, 2007, Mosby.)
Fig. 13-37 Schematic illustration of the histology of the thymus.
The functional thymus consists of two cell populations: the stromal cells and the thymocytes. The stromal cells include (1) the subcapsular epithelial cells also lining the trabeculae and perivascular spaces; (2) the cortical epithelial cells of ectodermal origin; (3) the medullary epithelial cells of endodermal origin that give rise to Hassall’s corpuscles; (4) macrophages present in both cortex and medulla, involved in the removal of apoptotic thymocytes eliminated during clonal selection; and (5) dendritic cells of bone marrow origin, confined to the medulla. TCR, T-cell receptor. (Adapted from Kierszenbaum AL: Histology and cell biology: an introduction to pathology, ed 2, St Louis, 2007, Mosby.)
The thymus attains its maximal mass relative to body weight at birth and involutes after sexual maturity. The lymphoid and epithelial components are gradually replaced by loose connective tissue and fat, although remnants remain histologically, even in aged animals.
Responses to injury
The responses of the thymus to injury are listed in Box 13-4, and the most common ones are lymphoid atrophy, inflammation, and neoplasia. The most common change is lymphoid atrophy caused by physical and physiologic stresses, toxins, drugs, and viral infections.
Atrophy: The causes of lymphoid atrophy are listed in Box 13-5. Because the thymus does not contain any lymphopoietic tissue, it depends on the bone marrow for the supply of T lymphocytes. Thus thymic lymphoid atrophy can be either the result of an inadequate supply of lymphocytes from the bone marrow or lysis of lymphocytes (lymphocytolysis) in the thymus.
Thymic atrophy must be differentiated from involution, which normally begins at sexual maturity. This distinction is difficult to make, unless the change is extreme or age-matched control animals are available. Viruses commonly cause injury to the thymus. Many viruses cause destruction of the lymphoid portion (e.g., from lymphocytolysis in equine herpesvirus-1 [EHV-1]) in aborted foals and feline panleukopenia virus in cats).
Inflammation: Inflammation of the thymus is rare. Inflammatory cells (neutrophils and sometimes macrophages) are often present within keratinized Hassall’s corpuscles during involution, and this should not be mistaken for a true thymitis. A diffuse infiltration of inflammatory cells into the thymus has also been reported in salmon poisoning disease of dogs (see Chapter 7) and a diffuse infiltration of macrophages in epizootic bovine abortion (see Chapter 18). Porcine circovirus 2, the etiologic agent of postweaning multisystem wasting syndrome (PMWS) in pigs, is unusual in that it causes a granulomatous inflammation in lymphoid organs, which in the thymus affects chiefly the medulla (see the section on Disorders of Pigs).
Thymic Enlargement: Thymic enlargement is rare and occasionally is the result of hemorrhage, hematomas, or tumors and is discussed further in the section on Disorders of Domestic Animals.
Neoplasia: Primary malignant tumors of the thymus are chiefly thymomas and lymphomas and are discussed further in the section on Disorders of Domestic Animals. Thymomas are formed from the epithelial component of the thymus, and usually with a benign proliferating lymphoid component. They are encapsulated, grow slowly, and rarely metastasize. Thymic lymphomas have a malignant proliferating lymphoid component and are most frequent in young cattle, dogs and cats. Histologically, they are poorly encapsulated and usually have an increased mitotic rate. Metastases can be widely disseminated, but are rare.
Spleen
Understanding the relationships between anatomic structures and the different functions of the spleen is complicated. There are also anatomic differences among domestic animal species and a confused terminology. Most of the latter is the result of the complicated vascular structure of the spleen and changes in terminology over the years. For example, “sinus” and “sinusoid” have both been applied to the “splenic sinusoids,” which in domestic animals are present only in the dog. Spleens, which have sinusoids, are often designated “sinusal” rather than “sinusoidal” and spleens without sinusoids as nonsinusal. Adding to the confusion, the word “sinus” has been used for the red pulp vascular spaces, which have also been called reticular spaces of the red pulp, interstices of the reticulum meshwork, and capillaries. Similarly, the white pulp has been termed splenic nodules, splenic follicles, and Malpighian corpuscles.
In this chapter, we use the following terms: sinusoid, red pulp vascular space, marginal sinus, marginal zone, periarteriolar lymphoid sheath (PALS), periarteriolar macrophage sheath (PAMS), splenic or lymphoid follicles, and splenic cords.
Structure
The spleen, suspended in the gastrosplenic ligament and covered by peritoneum, is located in the left cranial hypogastric region of the abdominal cavity between the diaphragm, stomach, and the body wall, except in domestic ruminants, where it is closely adhered to the left dorsolateral aspect of the rumen. The gross morphology (shape and size) of the spleen varies markedly among domestic animals, but generally it is a flattened, elongated organ. The spleen is covered by a thick fibromuscular capsule from which numerous intertwining fibromuscular trabeculae extend into the parenchyma and which along with the reticulum form the supporting stroma for the parenchymal components. The splenic parenchyma, unlike that of many organs, does not have a cortex and medulla but is divided into two distinct structural and functional components: the red pulp and white pulp. The red pulp consists of cells of the monocyte-macrophage system in the splenic cords and PAMS and red pulp vascular spaces and associated stroma (reticular cells and fibroblasts and myocytes in the trabeculae). The white pulp consists lymphocytes of the B lymphocyte regions (splenic follicles), T lymphocyte regions (PALS), and the marginal zone in which are macrophages, antigen-presenting cells, and trafficking B and T lymphocytes. There are minor differences in some of the splenic components of the domestic animals (e.g., the cat has a small marginal sinus but a well-developed PAMS).
The arterial and venous vasculature have minor differences among the domestic animals. The following account is correct for the canine spleen, and if the sinusoids, which are only in the canine spleen, are omitted, it applies in general to the nonsinusal spleens of the other domestic animals. The artery supplying the spleen is the splenic artery, a branch of the celiac artery, which is a major branch of the abdominal aorta. The splenic artery penetrates the splenic capsule at the hilus, branches, and enters the fibromuscular trabeculae as the trabecular artery (Fig. 13-38). As it leaves the trabecula, the trabecular artery becomes the central arteriole of the white pulp and is surrounded by a cuff of T lymphocytes (PALS). Adjacent to or eccentrically embedded in the PALS is a splenic follicle (B lymphocytes). A branch of the central artery goes to the marginal sinus, which surrounds the splenic follicle and carries blood-borne antigens and trafficking B and T lymphocytes into close contact with the follicle (Fig. 13-39). In the dog, the marginal sinus drains into the sinusoids, but in other domestic animals, it drains into the red pulp vascular spaces. It is significant that the first branch of the central artery goes to the marginal sinus. As a result, antigens, bacteria, particles, and other material are delivered to the marginal sinus first, before either the sinusoids (in the dog) or red pulp vascular spaces. After emerging from the PALS, the arteriole is surrounded by a sheath of macrophages known as PAMS. The PAMSs are very prominent in pigs. It then becomes the penicillary arteriole, and in horses, cattle, pigs, and cats, its terminal branches empty into the red pulp vascular space lined by reticular cells, which lie in and between the splenic cords. The term red pulp vascular spaces used here has the advantage of being descriptive and correct and clearly differentiates them from venous sinuses and sinusoids. In dogs and humans, branches of the central arteriole of the white pulp enter into venous sinusoids, which are lined by a discontinuous endothelium and these empty into splenic venules. The anatomic arrangement of arterioles opening into sinusoids is known as a closed circulation because the blood flow is through arterioles, capillaries, sinusoids, and venules, all of which are vessels lined by endothelium. The other route is through the red pulp vascular spaces, which are not lined by endothelium but by reticular cells and is designated an “open system” (see Fig. 13-38). Because the dog has sinusoids and red pulp vascular spaces, it has both open and closed splenic circulations. Blood flowing through either the sinusoids or red pulp vascular spaces is exposed to the surveillance of macrophages. These are located perivascularly around sinusoids, but their pseudopodia project into the sinusoidal lumen through the spaces in the discontinuous endothelium. In the red pulp, macrophages are present in the vascular spaces, attached to the reticular walls, and in the splenic cords. Blood then drains into the splenic venules, splenic veins, and ultimately the portal vein, which empties into the liver.
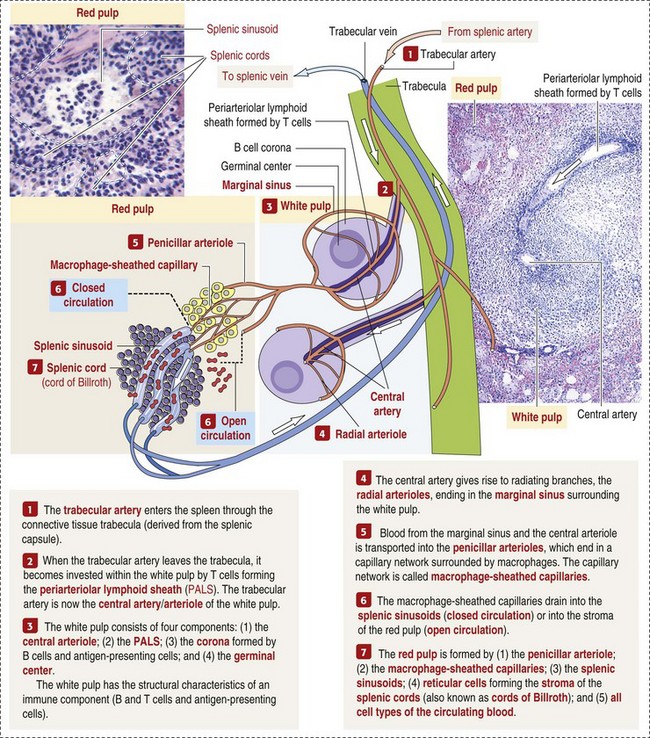
Fig. 13-38 Schematic representation of the human spleen.
The blood flow depicted is applicable in domestic animals only to the canine spleen, which has splenic sinusoids. The other domestic animals do not have splenic sinusoids. (Adapted from Kierszenbaum AL: Histology and cell biology: an introduction to pathology, ed 2, St Louis, 2007, Mosby.)
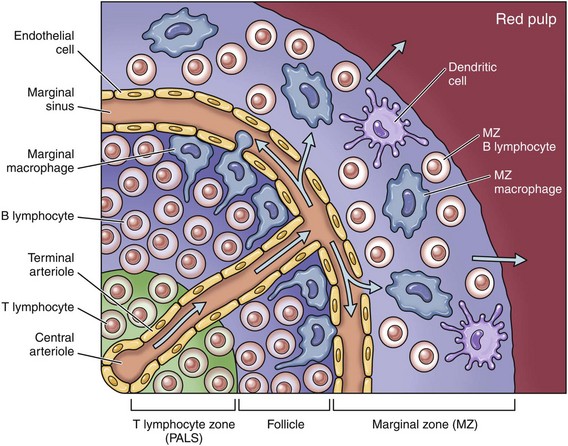
Fig. 13-39 Structure of the marginal zone in the splenic follicle.
Antigens from infections microorganisms enter the follicle via the central and terminal arterioles and reach the marginal sinus where they are phagocytosed by macrophages. Macrophages of the monocyte-macrophage system in the reticulum of the marginal zone, which is classified as part of the white pulp, form an important defense mechanism in the spleen through phagocytosis, antigen processing, and antigen presentation in splenic follicles to B lymphocytes resulting in the formation of immunoblasts that mature to plasma cells.