26 THE STRUCTURE AND FUNCTION OF THE DIGESTIVE SYSTEM
INTRODUCTION
The digestive system breaks down ingested food, prepares it for uptake by the body’s cells, provides body water and eliminates wastes. The system consists of (a) the gastrointestinal tract and (b) the accessory organs of digestion: the salivary glands, liver, gallbladder and pancreas. (The adjective gastric refers to the stomach; hence the name ‘gastrointestinal tract’ reflects its main components, the stomach and intestines.)
Food breakdown begins in the mouth with chewing and continues in the stomach, where food is churned and mixed with acid, mucus, enzymes and other secretions. From the stomach, the partially digested food passes into the small intestine, where biochemical agents and enzymes secreted by the liver and pancreas break it down into absorbable components of proteins, carbohydrates and fats. These nutrients are absorbed as they pass through the walls of the small intestine into blood vessels and lymphatics, which carry them to the liver for storage or further processing.
Ingested substances and secretions that are not absorbed in the small intestine pass into the large intestine, where fluid continues to be absorbed. Fluid wastes travel through the bloodstream to the kidneys and are eliminated in the urine. Solid wastes pass into the rectum and are eliminated from the body through the anus.
Except for chewing, swallowing and defecation of solid wastes, the movements of the digestive system (gastrointestinal motility) are all controlled by hormones and the autonomic nervous system. Autonomic innervation, both sympathetic and parasympathetic, is controlled by centres in the brain and by local stimuli that are mediated at plexuses (networks of nerve fibres) within the gastrointestinal walls.
AN OVERVIEW OF THE DIGESTIVE SYSTEM
The gastrointestinal tract and accessory organs
The gastrointestinal tract (also known as the alimentary canal, or gut) is essentially a long tube, extending from the mouth to the anus, which is divided into different organs (see Figure 26-1):
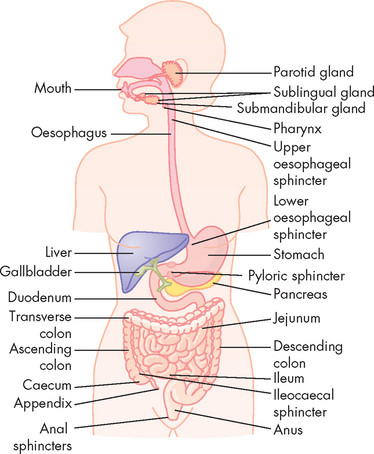
FIGURE 26-1 The gastrointestinal tract.
Source: Based on Guyton AC, Hall JE. Textbook of medical physiology. Philadelphia: Saunders; 11th edn.
The upper digestive tract consists of structures from the mouth to the stomach, while the intestines form the lower digestive tract. The specific structures and functions differ substantially at each organ. A number of sphincters are located between the different organs and these have an important role in controlling the passage of food. The sphincters of the gastrointestinal tract are as follows:
Sphincters are formed by a specialised ring of circular muscle — when contracted, food cannot progress through the sphincter.
A number of accessory organs are anatomically close to the gastrointestinal tract and these secrete substances that are vital for normal gastrointestinal function. The main accessory structures are the salivary glands, liver, gallbladder and pancreas. Sphincters are located where the accessory organs empty into the gastrointestinal tract and these must be relaxed to allow secretions to enter the gut.
The main digestive processes performed by the digestive system are:
Chemical digestion mainly occurs with the action of enzymes, which are molecules produced and secreted by the digestive system. The function of enzymes is to speed up chemical reactions, and in the digestive system they help to break food down quickly. Usually, enzyme names end in ‘-ase’, so this will help you to recognise them easily.
It is only when food is broken down into small components that it can be absorbed into the bloodstream. From here, the small molecules from the food are able to travel through the blood and become available for use by all body cells (see Figure 26-2). Throughout this chapter, you will learn the details of how this works.
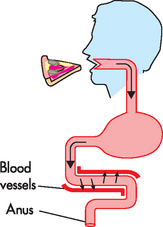
FIGURE 26-2 An overview of digestive system function.
After food is eaten, it travels through the digestive system where it is progressively broken down into smaller components. These small molecules must cross over the gut wall and into the nearby bloodstream, where they become available to body cells. Any molecules that are not absorbed exit the body in the faeces.
Histology of the gastrointestinal tract
The gastrointestinal tract consists of four distinct layers. From the lumen outwards, they are the mucosa, the submucosa, the muscularis and the serosa. These concentric layers vary in thickness and each layer has sublayers (see Figure 26-3).
 The mucosa consists of simple columnar epithelium, which lines the lumen and is therefore in direct contact with food. Scattered among this epithelial layer are goblet cells, which have the important function of secreting mucus that allows for lubrication during the passage of food. The proportion of goblet cells varies in some organs, depending on the requirement for mucus secretion. The lamina propria lies underneath the epithelium and consists of connective tissue; it provides the capillaries for the epithelial layer. A thin layer of smooth muscle called the muscularis mucosae provides the muscular activity for local movements. Some glands are also formed by the mucosal layer in some organs. The submucosa is essentially connective tissue and contains large blood vessels, nerves and secretory glands. The muscularis, as its name suggests, consists of muscle and has two sublayers:
The outermost layer of connective tissue is the serosa, which contains and supports the gastrointestinal tract.
The mucosa consists of simple columnar epithelium, which lines the lumen and is therefore in direct contact with food. Scattered among this epithelial layer are goblet cells, which have the important function of secreting mucus that allows for lubrication during the passage of food. The proportion of goblet cells varies in some organs, depending on the requirement for mucus secretion. The lamina propria lies underneath the epithelium and consists of connective tissue; it provides the capillaries for the epithelial layer. A thin layer of smooth muscle called the muscularis mucosae provides the muscular activity for local movements. Some glands are also formed by the mucosal layer in some organs. The submucosa is essentially connective tissue and contains large blood vessels, nerves and secretory glands. The muscularis, as its name suggests, consists of muscle and has two sublayers:
The outermost layer of connective tissue is the serosa, which contains and supports the gastrointestinal tract.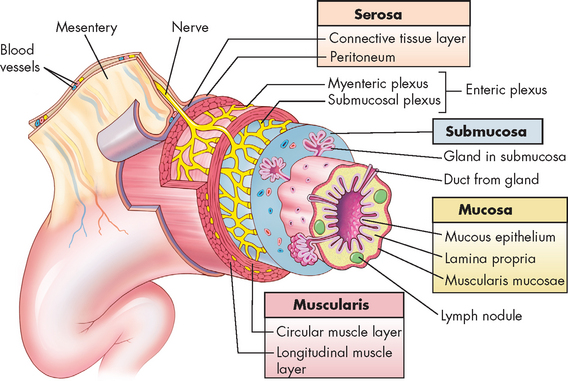
FIGURE 26-3 The wall of the gastrointestinal tract.
The wall of the gastrointestinal tract is made up of four layers with a network of nerves between the layers. This generalised diagram shows a segment of the gastrointestinal tract. Note that the serosa is continuous with a fold of serous membrane called a mesentery. Note also that digestive glands may empty their products into the lumen of the gastrointestinal tract by way of ducts.
Source: Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Nervous control of the digestive system
A network of intrinsic nerves called the enteric nervous system controls digestive function and is located solely within the layers of the digestive tract (see Figure 26-4). It consists of the following:
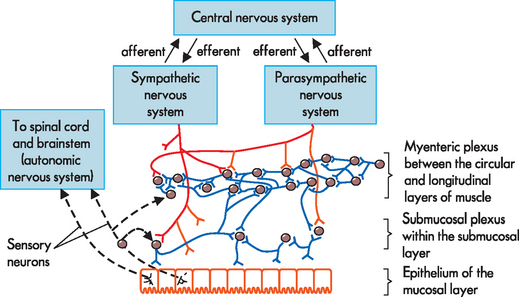
FIGURE 26-4 Neural control of the gut wall.
The main control is by the millions of neurons in the enteric nervous system, which also communicates with the sympathetic and parasympathetic nervous system branches. Afferent or sensory information travels through the gut wall towards the central nervous system, while efferent or motor information travels from the central nervous system towards the gut.
Interestingly, not only does the enteric nervous system control digestive activity using efferent signals (outflow from neurons to target cells), but there is also an important role for sensory neurons in this system. Specialised neurons are able to sense the local environment within the digestive system (such as within the contents of the stomach) and can relay afferent signals into the enteric nervous system. In particular, there are chemoreceptors to sense chemicals such as toxins in food, mechanoreceptors that can sense the mechanical forces of gut activity and stretch, and nociceptors to detect pain (refer to Chapter 6 for receptor types).1 On receiving information about the environment within the gut, the nervous system can alter function accordingly. For example, if food that contains a high amount of bacterial toxins reaches the intestines, the appropriate response may be diarrhoea to remove the toxins from the body. The intestinal contractions that cause diarrhoea require substantial amounts of contractions that must be initiated and coordinated by the nervous system. Also, sensory information from mechanoreceptors can detect when the stomach is stretched, which is vital in limiting further food intake temporarily.
In addition to the enteric nervous system, the gastrointestinal system is also controlled by the well-known branches of the autonomic nervous system: the parasympathetic and sympathetic divisions. Parasympathetic activity increases the function of the digestive system, such as increasing secretions and increasing motility, and causes relaxation of sphincters, thereby allowing food to progress through the system. Most parasympathetic innervation of the gastrointestinal system is via the vagus nerves (cranial nerve X). In contrast, the sympathetic nervous system, which coordinates the body’s response to stress (fight or flight response), decreases digestive processes and constricts sphincters. The sympathetic innervation of the digestive system is via the splanchnic nerves.
Motility
The majority of motility (or movement) in the digestive system is under involuntary control. Some voluntary control occurs at the mouth, pharynx and anus (and is discussed fully in the relevant sections). There are two types of motility:
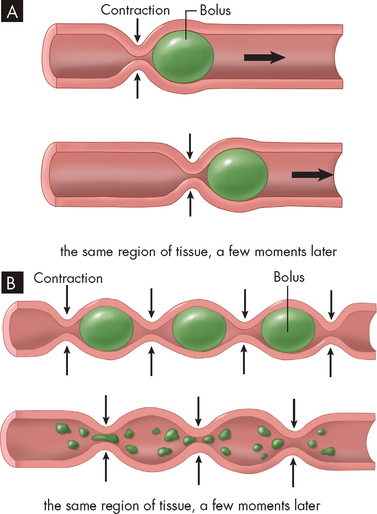
FIGURE 26-5 Peristalsis and segmentation.
A Peristalsis is a progressive type of movement in which food is propelled from point to point along the gastrointestinal tract. The ring of contraction moves like a wave along the tract to push the food forward. B Segmentation is a back-and-forth action that breaks apart chunks of food and mixes in digestive juices. Ring-like regions of contraction occur at intervals along the gastrointestinal tract. Previously contracted regions relax and adjacent regions contract, effectively ‘chopping’ the contents of each segment into smaller chunks, resulting in effective mixing.
Source: Based on Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Motility is controlled by a number of hormones, as well as the enteric nervous system.
Splanchnic blood flow
Splanchnic blood flow provides blood to the oesophagus, stomach, small and large intestines, liver, gallbladder, pancreas and spleen (see Figure 26-6). Blood flow is regulated by cardiac output and blood volume, the autonomic nervous system, hormones and local autoregulatory blood flow mechanisms. The splanchnic circulation also serves as an important reservoir of blood volume to return the blood to the heart when needed.
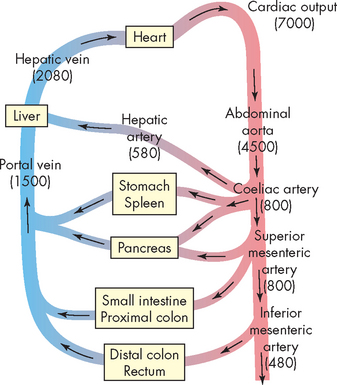
FIGURE 26-6 The major blood vessels and organs supplied with blood in the splanchnic circulation.
Numbers in parentheses reflect approximate blood flow values (mL/min) for each major vessel in an 80 kg resting adult human. Arrows indicate the direction of blood flow.
Source: Based on Johnson LR. Gastrointestinal pathophysiology. St Louis: Mosby; 2001.
The main nutrients
The energy from the food we eat comes from the three main nutrient groups: proteins, fats and carbohydrates. You will notice that these are the main molecules of the body that we examined in Chapter 1. These are the nutrients that will receive most of our attention in this chapter. Small amounts of vitamins (organic molecules) and minerals (inorganic molecules) are also essential for the human body and we briefly look at those as well.
THE MOUTH, PHARYNX AND OESOPHAGUS
Anatomy and physiology of the mouth, pharynx and oesophagus
The mouth (oral cavity or buccal cavity) is a reservoir for the chewing and mixing of food with saliva (see Figure 26-7). As food particles become smaller and move around in the mouth, the taste buds (or chemoreceptors) and olfactory nerves are continuously stimulated, adding to the enjoyment of eating. The tongue’s surface contains thousands of taste buds, which can distinguish salty, sour, bitter and sweet tastes. Food tastes and odours help to initiate salivation and the secretion of gastric juice in the stomach. There are 32 permanent teeth in the adult mouth and they are important for speech and mastication (chewing).
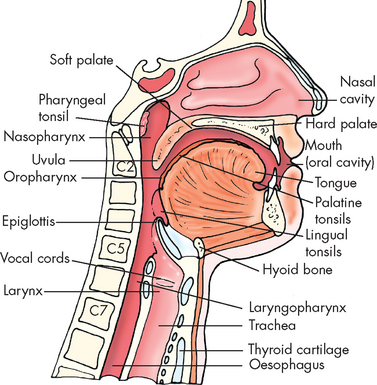
FIGURE 26-7 Structures associated with the mouth, pharynx and oesophagus.
Source: Based on Frank ED, Long BW, Smith BJ. Merrill’s atlas of radiographic positioning and procedures. Volume 2. 11th edn. St Louis: Mosby; 2007.
The pharynx is the region posterior to the mouth and nose and leads into the oesophagus. It consists of three regions: nasopharynx, oropharynx and laryngopharynx (see Figure 26-7). The oesophagus is a hollow, muscular tube approximately 25 cm long that conducts substances from the oropharynx, through the thoracic cavity and into the stomach. Each end of the oesophagus is opened and closed by a sphincter. The upper oesophageal sphincter keeps air from entering the oesophagus during respiration; and the lower oesophageal sphincter (also known as the cardiac sphincter or gastro-oesophageal sphincter) prevents regurgitation from the stomach back into the oesophagus.
Salivation
There are three pairs of salivary glands: the left and right submandibular, sublingual and parotid glands (see Figure 26-8). These glands secrete approximately 1.5 L of saliva into the mouth per day, via a duct with the same name as the gland itself (for example, the parotid gland empties into the mouth via the parotid duct). Saliva consists mostly of water with mucus, as well as ions including sodium, bicarbonate, chloride and potassium. Another important component of saliva, salivary amylase, is discussed later in ‘Digestion in the mouth, pharynx and oesophagus’ below.
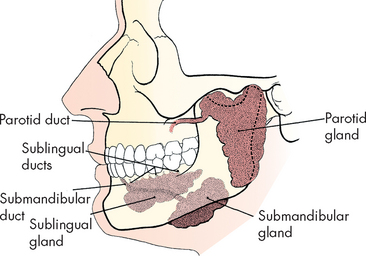
FIGURE 26-8 Salivary glands from the left lateral aspect.
Source: Frank ED, Long BW, Smith BJ. Merrill’s atlas of radiographic positioning and procedures. Volume 2. 11th edn. St Louis: Mosby; 2007.
The composition of saliva depends on the rate of secretion. The bicarbonate concentration of saliva sustains a pH of about 7.4, which neutralises bacterial acids and prevents tooth decay. Saliva also contains antimicrobial substances (such as lysosyme and immunoglobulin A, IgA), which help prevent infection. Exogenous fluoride (namely, fluoride in drinking water) is also secreted in the saliva, providing additional protection against tooth decay.
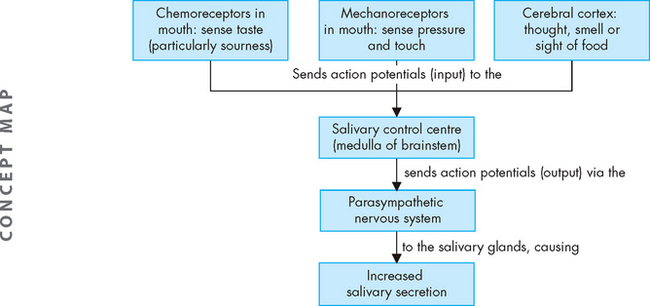
Saliva is secreted at a relatively constant rate, keeping the mouth moist and facilitating speech and swallowing. The amount of saliva is increased in response to nervous system activity. You will recall that control centres for vital body functions are located in the brainstem (such as the cardiac and respiratory control centres) — this is also true for the salivary control centre, which is located in the medulla. It is the parasympathetic nervous system that is mainly responsible for increased secretions (see Figure 26-9). Sensory receptors in the mouth sense the presence of particular tastes (such as sour or acid), and mechanoreceptors sense the physical presence of objects in the mouth. Interestingly, the pressure receptors are activated whether the object in the mouth is food or not — for example, if you continue sucking the stick from an ice-block after the ice-block has been eaten, the mechanoreceptors will continue to be activated. Inputs (action potentials) from these sensory neurons reach the salivary control centre in the medulla, which results in an increase in salivary secretions (mediated by outputs from the parasympathetic nervous system). The higher centres of the brain can also stimulate secretion of saliva, such as the conscious thought of food in the cerebral cortex — this too results in action potentials reaching the salivary control centre and increased salivation.
Swallowing
Swallowing (or deglutition) occurs when food mixed with saliva, now known as a bolus, moves onwards from the mouth. The upper third of the oesophagus contains striated muscle (voluntary) that is directly innervated by motor neurons. The lower two-thirds contain smooth muscle (involuntary) that is innervated by the parasympathetic nervous system. Peristalsis is stimulated when afferent fibres distributed along the length of the oesophagus sense changes in wall tension caused by stretching as food passes. The greater the tension, the greater the intensity of oesophageal contraction. Occasionally, intense contractions cause pain similar to ‘heartburn’ or angina. (Heartburn is chest pain associated with the oesophagus.)
Swallowing is coordinated primarily by the swallowing control centre in the medulla. During the oropharyngeal (voluntary) phase, the following steps occur (see Figure 26-10A–C):
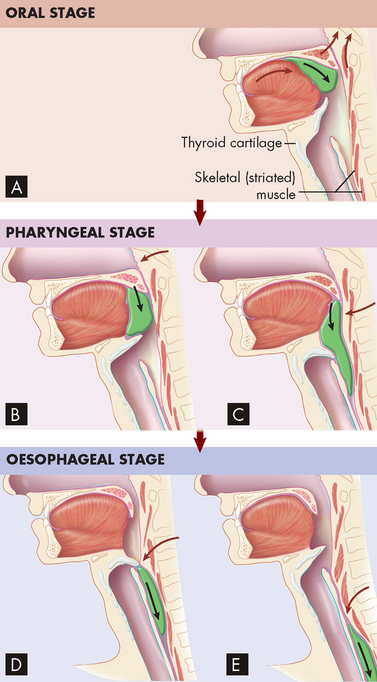
FIGURE 26-10 Deglutition (swallowing).
A–C Oropharyngeal stage. A During this stage of swallowing, a bolus of food is voluntarily formed on the tongue and pushed against the palate and into the oropharynx. Notice that the soft palate and uvula prevent food from entering the nasopharynx. B After the bolus enters the oropharynx, involuntary reflexes push the bolus down towards the oesophagus. C The upward movement of the larynx and downward movement of the bolus close the epiglottis and thus prevent food from entering the lower respiratory tract. D–E Oesophageal stage. D Involuntary reflexes of skeletal and smooth muscle in the wall of the oesophagus move the bolus through the oesophagus towards the stomach. E After the bolus has passed, the epiglottis moves upwards, opening the larynx and trachea to respiration.
Source: Based on Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
This entire sequence of moving food from the mouth through the pharynx takes place in less than 1 second.
The oesophageal phase of swallowing proceeds as follows (see Figure 26-10D–E):
This phase takes about 5–10 seconds because the lower oesophageal sphincter is normally constricted and serves as a barrier between the stomach and oesophagus.
Peristalsis that immediately follows the oropharyngeal phase of swallowing is called primary peristalsis. If a bolus becomes stuck in the oesophagus, secondary peristalsis — a wave of contraction and relaxation independent of voluntary swallowing — occurs. This is in response to stretch receptors in the oesophagus, which increase impulses from the swallowing centre of the brain.
Digestion in the mouth, pharynx and oesophagus
The mouth is the first site of mechanical digestion, whereby the chewing action of the teeth causes physical breakdown of food into smaller particles. Once the food is small enough (and has been adequately moistened with saliva), it is ready to be swallowed. There is no further mechanical digestion in the pharynx or oesophagus.
Saliva contains salivary amylase, which is an enzyme that initiates carbohydrate digestion — this is the only enzyme involved in chemical digestion in the mouth. Carbohydrate digestion by salivary amylase may continue when the bolus (mixed with enzyme) continues into the stomach. After the enzyme mixes with the acidic environment of the stomach, it becomes inactivated.
None of the main nutrient groups (carbohydrates, proteins and fats) are digested sufficiently to be absorbed into the bloodstream from the mouth, pharynx or oesophagus. However, some drugs are administered in the mouth to be absorbed directly into the bloodstream from salivary glands, as this allows the drugs to enter the bloodstream quickly. Examples include glyceryl trinitrate (for the treatment of angina pectoris, discussed previously in Chapter 23) and fetanyl (for the immediate treatment of cancer pain).
THE STOMACH
Anatomy and physiology of the stomach
The stomach is a muscular organ just inferior to the diaphragm that stores food during eating, secretes digestive juices, mixes food with these juices and propels partially digested food, now called chyme, into the small intestine. The anatomy of the stomach is shown in Figure 26-11. Its major anatomical boundaries are the lower oesophageal sphincter, where food passes into the stomach; the greater and lesser curvatures; and the pyloric sphincter, which relaxes as food is propelled through the pylorus (gastroduodenal junction) into the duodenum. Functional areas are the fundus (upper portion), body (middle portion) and antrum (lower portion).
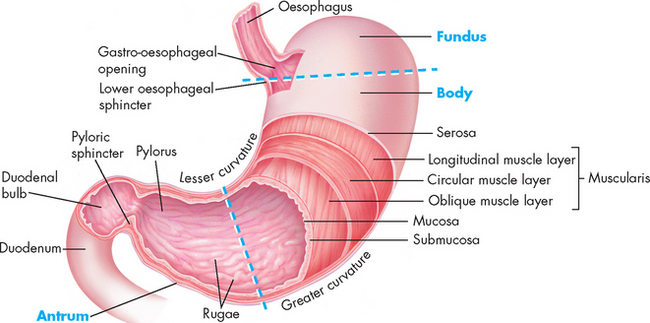
A portion of the anterior wall has been cut away to reveal the muscle layers of the stomach wall. Note that the mucosa lining the stomach forms folds called rugae. The dotted lines distinguish the fundus, body and antrum of the stomach.
Source: Based on Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
The muscularis layer has an inner circular muscle layer and an outer longitudinal layer, as described earlier. An additional sublayer of oblique muscle occurs in the stomach and it is the most prominent, lying between the submucosa and the circular muscle layer.
Gastric motility
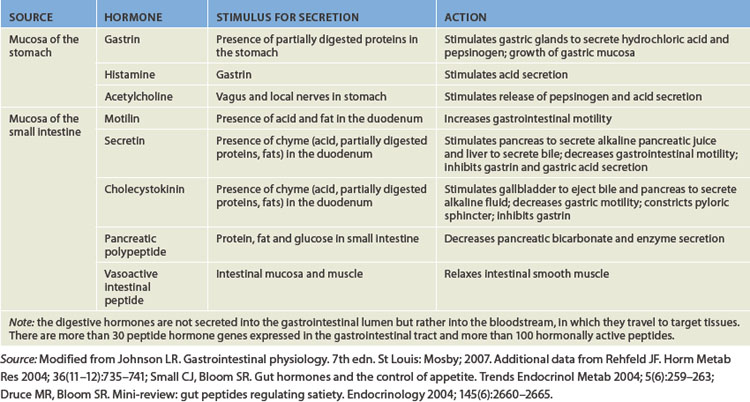
In the resting state, the stomach is small and contains about only 50 mL of fluid. However, the stomach has a tremendous capacity and can expand to hold up to 4 L. Swallowing causes the fundus to relax to receive a bolus from the oesophagus. Relaxation is facilitated by gastrin and cholecystokinin — two polypeptide hormones secreted by the gastrointestinal mucosa (the actions of digestive hormones are summarised in Table 26-1).
Gastric motility increases with the initiation of peristaltic waves, which sweep over the body of the stomach towards the antrum. The rate of peristaltic contractions is approximately 3 per minute and is influenced by neural and hormonal activity. Gastrin, motilin (an intestinal hormone) and the activation of the parasympathetic nervous system (travelling via the vagus nerve) increase contraction. Sympathetic activity and secretin (another intestinal hormone) are inhibitory and decrease the rate of contractions. The rate of peristalsis is mediated by pacemaker cells that initiate a wave of depolarisation (basic electrical rhythm), which moves from the upper part of the stomach to the pylorus.
The mixing and emptying of food from the stomach takes several hours. Mixing occurs as food is propelled towards the antrum. As food approaches the pylorus, the velocity of the peristaltic wave increases, forcing the contents back towards the body of the stomach. This retropulsion effectively mixes food with digestive juices and the oscillating motion breaks down large food particles. With each peristaltic wave, a small portion of the gastric contents (chyme) passes through the pylorus and into the duodenum, as the pyloric sphincter opens only slightly to regulate the rate of entry into the duodenum (see Figure 26-12).
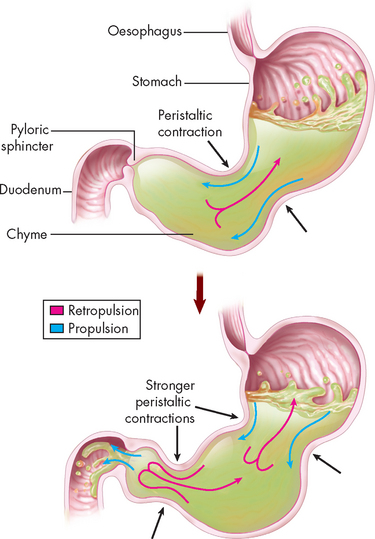
FIGURE 26-12 Stomach mechanical events.
Mixing actions in the stomach include both propulsion (forward movement) and retropulsion (backward movement). As peristaltic contractions become stronger, some of the liquid chyme squirts past the pyloric sphincter (which has decreased its muscle tone) and into the duodenum. The stomach continues to mix the chyme as it is gradually released into the duodenum.
Source: Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
The rate of gastric emptying (movement of gastric contents into the duodenum) depends on the volume, osmotic pressure and chemical composition of the gastric contents. Larger volumes of food increase gastric pressure, peristalsis and rate of emptying. Food remains in the stomach for up to 4 hours, although a further 2 hours may be required for fatty meals.2 Products of fat digestion, which are formed in the duodenum by the action of bile from the liver and enzymes from the pancreas, stimulate the secretion of cholecystokinin. This hormone inhibits gastric motility and decreases gastric emptying so that fats are not emptied into the duodenum at a rate that exceeds the rate of bile and enzyme secretion. Osmoreceptors in the wall of the duodenum are sensitive to the osmotic pressure of duodenal contents. The arrival of hypertonic or hypotonic gastric contents activates the osmoreceptors, which delay gastric emptying so that the duodenum can receive the contents. (This is why sports drinks are often formulated to be ‘isotonic’. Review ‘tonicity’ in Chapter 3.) The rate at which acid enters the duodenum also influences gastric emptying. Secretions from the pancreas, liver and duodenal mucosa neutralise gastric acid in the duodenum. The rate of emptying is adjusted to the duodenum’s ability to neutralise the incoming acidity.2
Gastric secretion
Stimulated by eating, the stomach secretes large volumes of gastric juices (or gastric secretions) directly into the lumen. These include mucus, acid, enzymes, intrinsic factor and gastroferrin. Intrinsic factor is necessary for subsequent intestinal absorption of vitamin B12 and gastroferrin facilitates small intestinal absorption of iron. The stomach also secretes some hormones into the bloodstream, including gastrin and histamine, which stimulate the gastric glands.
In the fundus and body of the stomach, the gastric glands of the mucosa are the primary secretory units (see Figure 26-13A). From the stomach lumen down to the depths of the glands, the cell types are the mucous neck cells, chief cells, parietal cells and finally the enteroendocrine cells (which secrete the hormones into the bloodstream). The rate of gastric secretions varies with the time of day. Generally, the rate and volume of secretion are lowest in the morning and highest in the afternoon and evening.
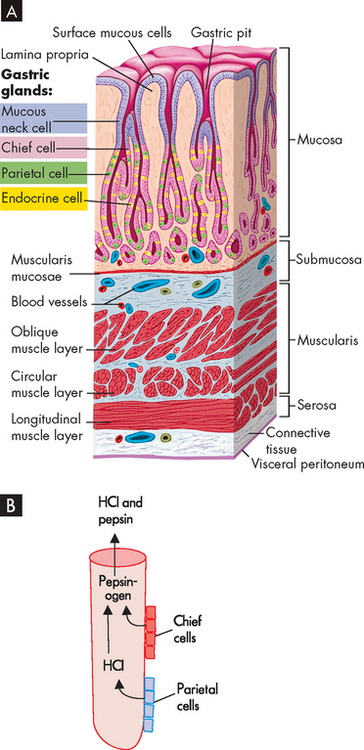
FIGURE 26-13 A Gastric pits and gastric glands. Gastric pits are depressions in the epithelial lining of the stomach. At the bottom of each pit is one or more tubular gastric glands. Chief cells produce the enzymes of gastric juice (such as pepsinogen), parietal cells produce stomach acid, and G cells produce the hormone gastrin. B Pepsin activation. Pepsinogen secreted by chief cells becomes activated to pepsin in the presence of hydrochloric acid (HCl).
Source: Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Gastric secretions are increased by parasympathetic nervous system activity, while the sympathetic nervous system decreases secretions. Unpleasant odours and tastes or strong emotions alter the relative sympathetic and parasympathetic nervous system control and can therefore alter secretions.
Acid
A high concentration of hydrochloric acid allows the stomach pH to be as low as 2 or 3 (a low pH is due to a high amount of acid). The major functions of gastric acid are to dissolve food fibres, act as a bactericide (kill bacteria) against swallowed microorganisms and convert pepsinogen to pepsin. The production of acid is by the parietal cells of the gastric glands.
Pepsin
Acetylcholine, gastrin and secretin stimulate the chief cells to release pepsinogen during eating. Pepsinogen is quickly converted to pepsin in the acid gastric environment (see Figure 26-13B). Pepsin is a proteolytic enzyme — that is, it breaks down protein into smaller components known as polypeptides in the stomach. Once chyme enters the duodenum, the alkaline environment of the duodenum inactivates pepsin.
Mucus
The gastric mucosa is protected from the digestive actions of acid and pepsin by a coating of alkaline (bicarbonate-rich) mucus called the mucosal barrier. Mucus is produced in large quantities in the stomach by the mucus neck cells of the gastric glands. The quality and quantity of mucus and the tight junctions between epithelial cells make gastric mucosa relatively impermeable to acid. Prostaglandins protect the mucosal barrier by stimulating the secretion of mucus and bicarbonate and by inhibiting the secretion of acid. Mucosal blood flow is important to maintaining mucosal protective functions. If epithelial cells lining the stomach are damaged, they are replaced quickly to protect underlying tissues.
Phases of gastric secretion
The secretion of gastric juice is influenced by numerous stimuli that together facilitate the process of digestion. There are three phases of gastric secretion: the cephalic phase, the gastric phase and the intestinal phase. Briefly, the cephalic phase actually occurs before food enters the stomach. The phase is activated by the thought, smell, sight and taste of food. These sensory inputs travel to the hypothalamus and then the medulla, which stimulates the stomach via the vagus nerve. The gastric phase is initiated once food enters the stomach, as it causes distension or stretch of the stomach. Most of the gastric juices are secreted in this phase. The hormone gastrin is released, due to chemoreceptor stimulation from the chyme, and this stimulates parietal cells to release hydrochloric acid. This lowers the pH and protein digestion occurs. The last phase, the intestinal phase, occurs when chyme starts to enter the duodenum, which causes the release of more acid and also triggers the release of secretin and cholecystokinin, hormones that chemically prepare the duodenum.
Digestion in the stomach
The main digestive process in the stomach is mechanical digestion by the strong muscular layers, whereby larger food particles are ground and churned into smaller particles. Chemical digestion also occurs, whereby hydrochloric acid and the enzyme pepsin break down proteins into smaller molecules.
Absorption from the stomach
Absorption from the stomach is limited: none of the main nutrients are absorbed here. However, both alcohol and aspirin can easily pass through the stomach’s mucosal lining and into the bloodstream. Excessive amounts of these substances can be irritating to the stomach mucosa and therefore it is advisable to have food in the stomach at the same time that alcohol or aspirin (and other anti-inflammatory drugs) are consumed, to help protect the mucus lining.
THE SMALL INTESTINE
Anatomy and physiology of the small intestine
The small intestine (or small bowel) is named ‘small’ due to its relatively small diameter of approximately 2.5 cm. It is about 5 m long and is functionally divided into three segments: the duodenum, the jejunum and the ileum (see Figures 26-1 and 26-14). These structures are not grossly different, but the jejunum has a slightly larger lumen. At the distal end, the ileocaecal valve, or sphincter, controls the flow of digested material from the ileum into the large intestine and prevents reflux into the small intestine.3
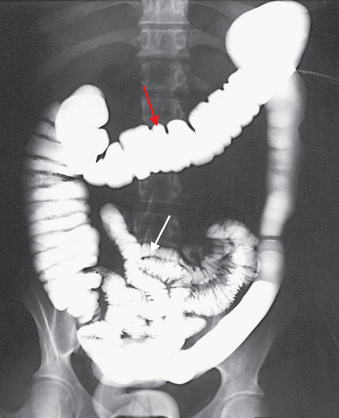
The proximal part of the small intestine (white arrow) and all of the large intestine (red arrow) are shown on this image.
Source: Frank ED, Long BW, Smith BJ. Merrill’s atlas of radiographic positioning and procedures. Volume 2. 11th edn. St Louis: Mosby; 2007.
The peritoneum is the serous membrane surrounding the organs of the abdomen and pelvic cavity. It is analogous to the pericardium and pleura, which surround the heart and lungs, respectively. The visceral peritoneum lies over the organs and the parietal peritoneum lines the wall of the abdominal cavity. The space between these two layers is called the peritoneal cavity and normally contains just enough fluid to lubricate the two layers and prevent friction during organ movement.
Associated with the anterior part of the peritoneal membranes is the omenta (plural; omentum = singular). The greater omentum drapes from the inferior part of the stomach over the intestinal fold and contains a considerable amount of fat. As fat builds up around the waist area, an increase in storage of fat within the greater omentum occurs. The lesser omentum is a much smaller membrane, located between the liver and stomach. We consider the storage of fat in greater detail in Chapter 35.
The duodenum lies behind the peritoneum (retroperitoneal) and is attached to the posterior abdominal wall. The ileum and jejunum are suspended in loose folds from the posterior abdominal wall by a peritoneal membrane called the mesentery. The mesentery facilitates intestinal motility and supports blood vessels, nerves and lymphatics.
There are important structural specialisations of the small intestine that force food to move through quite slowly — this is important as a large amount of digestion and absorption occurs here. Mucosal folds within the small intestine slow the passage of food, thereby providing more time for digestion and absorption (see Figure 26-15). The folds are most numerous and prominent in the jejunum and upper ileum.
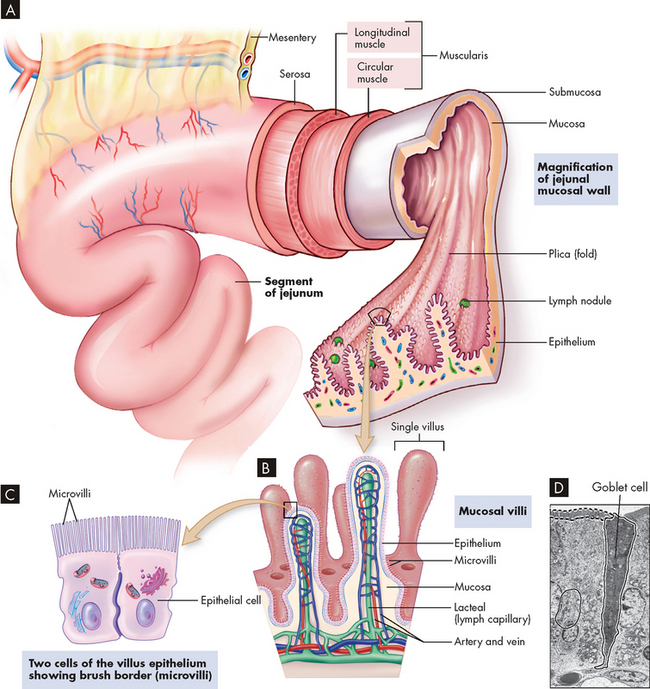
FIGURE 26-15 The wall of the small intestine.
A The folds of mucosa are covered with villi. B Each villus is covered with epithelium. C The epithelial cells have microvilli, which increase the surface area for absorption of food. D A microscope image of the epithelial cells of the small intestine, showing the microvilli. Note that a goblet cell containing mucus can also be seen.
Source: A, B & C Telser AG, Young, JK, Baldwin, KM. Elsevier’s integrated histology. St Louis: Mosby; 2007. D Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Absorption occurs through villi (plural; villus = singular), which cover the mucosal folds and are the functional units of the intestine. Each villus (see Figure 26-15A–B) secretes some of the enzymes necessary for digestion and absorbs nutrients. A villus is composed of absorptive columnar cells and mucus-secreting goblet cells of the mucosal epithelium. Near the surface, columnar cells closely adhere to each other at sites called tight junctions (literally, close together). Water and electrolytes are absorbed through these intercellular spaces. The surface of each columnar epithelial cell contains tiny projections called microvilli (see Figure 26-15C–D). Together the microvilli create a mucosal surface known as the brush border. The villi and microvilli greatly increase the surface area available for absorption. Coating the brush border is an ‘unstirred’ layer of fluid that is important for the absorption of substances other than water and electrolytes. The lamina propria (a connective tissue layer of the mucous membrane) lies beneath the epithelial cells of the villi and contains lymphocytes and macrophages, involved in inflammation and immune functions.
Central arterioles ascend within each villus and branch into a capillary array that extends around the base of the columnar cells and cascades down to the venules that lead to the portal circulation (see Figure 26-15B). A central lacteal, or lymphatic channel is also contained within each villus and is important for the absorption and transport of fat molecules. Contents of the lacteals flow to regional nodes and channels that eventually drain into the thoracic duct.4
The entire epithelial population is replaced about every 4–7 days. Many factors can influence this process of cellular proliferation. Starvation, vitamin B12 deficiency and chemotherapy (cytotoxic drugs) or irradiation suppress cell division and shorten the villi. The decreased absorption that results can cause diarrhoea and malnutrition.
Intestinal motility
The movements of the small intestine facilitate both digestion and absorption. Chyme coming from the stomach stimulates intestinal movements that mix in secretions from the liver, pancreas and intestinal glands. A churning motion brings the luminal contents into contact with the absorbing cells of the villi. The propulsive movements of peristalsis then advance the chyme towards the large intestine.
Neural reflexes along the length of the small intestine facilitate motility, digestion and absorption. The ileogastric reflex inhibits gastric motility when the ileum becomes distended. This prevents the continued movement of chyme into an already distended intestine. The intestinointestinal reflex inhibits intestinal motility when one part of the intestine is overdistended. Both of these reflexes require extrinsic innervation. The gastroileal reflex, which is activated by an increase in gastric motility and secretion, stimulates an increase in ileal motility and relaxation of the ileocaecal sphincter. This empties the ileum and prepares it to receive more chyme.
During prolonged fasting or between meals, particularly overnight, slow waves sweep along the entire length of the intestinal tract from the stomach to the terminal ileum. This action appears to propel residual gastric and intestinal contents into the colon.
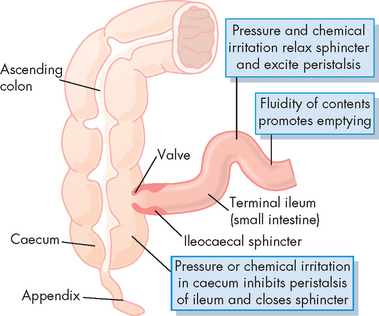
The ileocaecal sphincter marks the junction between the terminal ileum and the large intestine. This valve is intrinsically regulated and is normally closed. The arrival of peristaltic waves from the last few centimetres of the ileum causes the ileocaecal valve to open, allowing a small amount of chyme to pass through. Distention of the upper large intestine causes the sphincter to constrict, preventing further distension, as well as avoiding retrograde flow of intestinal contents (see Figure 26-16).
Digestion in the small intestine
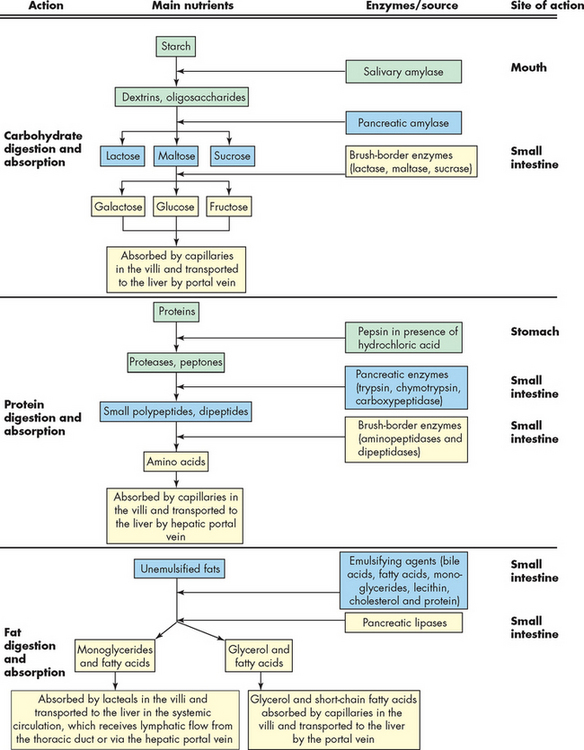
The chyme that passes into the duodenum is a liquid with small particles of undigested food. Importantly, the secretions from the accessory organs — the liver, gallbladder and pancreas — all empty into the duodenum. Digestion continues in the proximal portion of the small intestine by the action of pancreatic enzymes, intestinal enzymes and bile salts. There carbohydrates are broken down to monosaccharides and disaccharides; proteins are degraded further to amino acids and peptides; and fats are emulsified and reduced to fatty acids and monoglycerides (see Figure 26-17).
Absorption from the small intestine
The majority of all absorption occurs in the small intestine. The small molecules of the main nutrient groups (simple sugars, amino acids and fatty acids), along with water, vitamins and electrolytes, are absorbed across the intestinal mucosa by active transport, diffusion or facilitated diffusion. Products of carbohydrate, protein and lipid breakdown move into capillaries (of the villi) and then travel to the liver through the hepatic portal vein. Some types of digested fats cross into the lacteals and therefore travel through the lymphatic system and do not go through the liver. The lymphatic system empties into the systemic circulation near the heart, so here those lipids reach the bloodstream. Intestinal motility exposes nutrients to a large mucosal surface area by mixing chyme and moving it through the lumen. Different segments of the gastrointestinal tract absorb different nutrients. Sites of absorption are shown in Figure 26-18. Box 26-1 outlines the major processes involved in nutrient absorption.
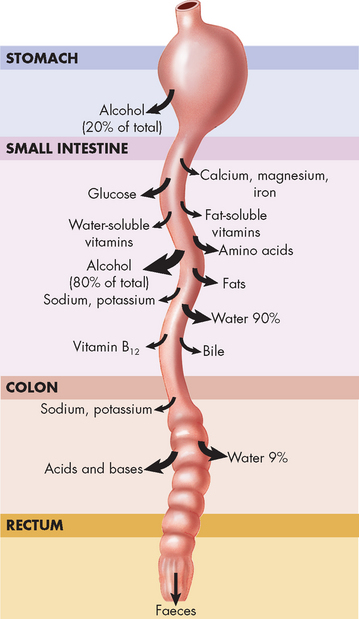
FIGURE 26-18 Absorption sites in the digestive tract.
The size of the arrow at each site indicates the relative amount of absorption of a particular substance at that site. Notice that most absorption occurs in the intestines, particularly the small intestine.
Source: Based on Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Box 26-1 MAJOR NUTRIENTS ABSORBED IN THE SMALL INTESTINE
Water and electrolytes
Carbohydrates
Proteins
Fats
ACCESSORY ORGANS OF DIGESTION
The liver, gallbladder and pancreas all secrete substances necessary for the digestion of chyme. These secretions are delivered to the duodenum through ducts. The liver produces bile, which contains salts necessary for fat digestion and absorption. Between meals, bile is stored in the gallbladder. The pancreas produces (1) enzymes needed for the complete digestion of carbohydrates, proteins and fats, and (2) an alkaline fluid that neutralises chyme, creating a duodenal pH that supports enzymatic action.
The liver
The liver weighs 1200–1600 grams. It is located under the right side of the diaphragm and is divided into right and left lobes (see Figure 26-19). The falciform ligament separates the right and left lobes and attaches the liver to the anterior abdominal wall. The round ligament (ligamentum teres) extends along the free edge of the falciform ligament, extending from the umbilicus to the inferior surface of the liver. The coronary ligament branches from the falciform ligament and extends over the superior surface of the right and left lobes, binding the liver to the inferior surface of the diaphragm. The liver is covered by the Glisson capsule, which contains blood vessels, lymphatics and nerves. When the liver is diseased or swollen, distension of the capsule causes pain, and the lymphatics may ooze fluid into the peritoneal space.
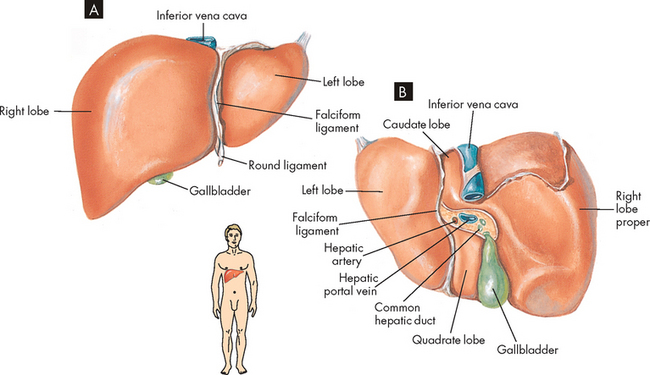
FIGURE 26-19 Gross structure of the liver.
A Anterior view. B Inferior view.
Source: Based on Thibodeau GA, Patton KT. Anatomy & physiology. 6th edn. St Louis: Mosby; 2007.
The liver’s blood supply is unusual, in that there are two inward sources (see Figure 26-20):
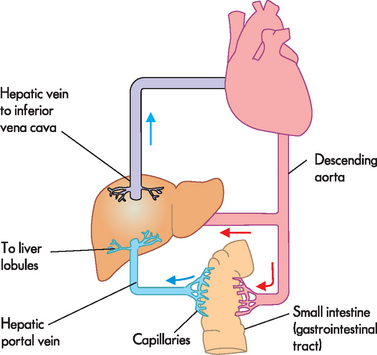
FIGURE 26-20 Hepatic portal circulation.
Substances are absorbed from the digestive tract into capillaries, which lead into the hepatic portal vein. This nutrient-rich blood enters the liver to be filtered. The other blood supply to the liver is via the hepatic artery, bringing oxygenated blood from the heart. All blood empties from the liver into the hepatic vein, which drains to the vena cava. Note that some small fatty molecules are absorbed into the lacteals to reach the lymphatic system, which does not travel through the hepatic portal circulation. Instead, the contents of lymphatic ducts empty directly into the bloodstream near the vena cava.
Within the liver lobes are multiple, smaller anatomical units called liver lobules (see Figure 26-21). They are formed of cords or plates of hepatocytes (literally ‘liver cells’), which are the functional cells of the liver. These cells can regenerate. In fact, an individual can survive even if more than half of the liver is removed, as the liver will grow back to its original size. Small capillaries, or sinusoids (leaky capillaries), are located between the plates of hepatocytes. They receive a mixture of blood from branches of the hepatic artery and hepatic portal vein. The liver has a pivotal function in processing the nutrient-rich blood from the digestive tract and therefore the hepatocytes have very rich access to the blood from these sinusoids. Blood from the sinusoids drains to a central vein in the middle of each liver lobule. Venous blood from all the lobules then flows into the hepatic vein, which empties blood away from the liver into the inferior vena cava.
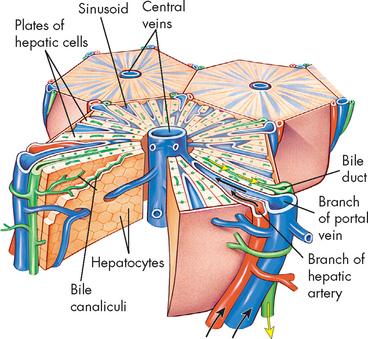
FIGURE 26-21 Diagrammatic representation of a liver lobule.
A central vein is located in the centre of the lobule with plates of hepatic cells disposed radially. Branches of the portal vein and hepatic artery are located on the periphery of the lobule, and blood from both perfuses the sinusoids. Peripherally located bile ducts drain the bile canaliculi that run between the hepatocytes.
Source: Based on Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
Small channels (bile canaliculi) conduct bile, which is produced by the hepatocytes, outwards to the bile ducts and eventually drain into the common bile duct (see Figure 26-21). This duct empties bile into the duodenum through an opening called the major duodenal papilla (sphincter of Oddi; see Figure 26-22).
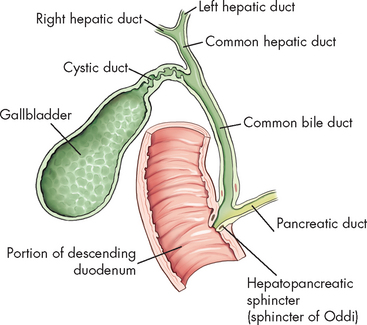
FIGURE 26-22 Gallbladder and extrahepatic biliary ducts.
Source: Bontrager KL, Lampignano, JP. Textbook of radiographic positioning and related anatomy. 7th edn. St Louis: Mosby; 2010.
The sinusoids of the liver lobules are lined with highly permeable endothelium. This permeability enhances the transport of nutrients from the sinusoids into the hepatocytes, where they are metabolised. The sinusoids are also lined with phagocytic Kupffer cells (tissue macrophages), which are part of the mononuclear phagocyte system. They remove foreign substances from the blood and trap bacteria. Between the endothelial lining of the sinusoid and the hepatocyte is the Disse space, which drains interstitial fluid into the hepatic lymph system.
Secretion of bile
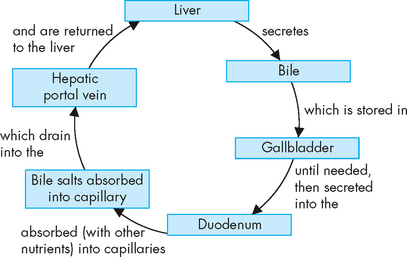
The liver hepatocytes assist intestinal digestion by producing and secreting 700–1200 mL of bile per day. Bile is an alkaline (bicarbonate-rich), bitter-tasting, yellowish green fluid that contains bile salts, cholesterol, bilirubin, electrolytes and water. It is formed by hepatocytes and secreted into the canaliculi. Bile salts, which are produced from cholesterol, are required for the intestinal emulsification of fats into small spheres known as micelles. These micelles can then be absorbed into the lacteals of the lymphatic system and then into the systemic blood circulation.
Having facilitated fat emulsification and absorption, most bile salts are actively absorbed in the terminal ileum and returned to the liver through the portal circulation for resecretion. The recycling of bile salts is termed the enterohepatic circulation (see Figure 26-23). The cycle of hepatic secretion, intestinal absorption and hepatic resecretion of bile acids completes the enterohepatic circulation.
Vascular and haematological functions
Because of its extensive vascular network, the liver can store a large volume of blood. The amount stored at any one time depends on pressure relationships in the arteries and veins. The liver also can release blood to maintain systemic circulatory volume in the event of haemorrhage.
Kupffer cells in the sinusoids of the liver remove bacteria and foreign particles from the hepatic portal blood. Because the liver receives all the venous blood from the gastrointestinal tract, Kupffer cells play an important role in destroying bacteria and preventing infections from the intestines reaching the rest of the body.
The liver also has haemostatic functions, in that it produces clotting factors necessary for coagulation — namely prothrombin, fibrinogen and clotting factors I, II, VII, IX and X (see Chapter 16). Vitamin K is a fat-soluble vitamin essential for the production of clotting factors. Because bile salts are needed for the absorption of fats, vitamin K absorption depends on adequate bile production in the liver. In this way, normal liver function is necessary for adequate absorption of vitamin K to allow adequate circulating levels of clotting factors.
Metabolism of nutrients
Fats
Some fatty molecules are absorbed into the lacteals and carried via the lymphatic system and then emptied directly into the bloodstream (near the heart) to travel around the circulation. Almost all substances other than fats, including proteins, carbohydrates, water, vitamins, minerals, drugs and contaminants, are absorbed from the small intestine to the blood, which is processed by the liver before reaching the rest of the circulation.
After being absorbed by lacteals in the intestinal villi, the fats enter the liver through the lymphatics, primarily as triglycerides. These can be used to produce metabolic energy (adenosine triphosphate, ATP) or they can be released into the bloodstream as lipoproteins (lipids bound to proteins). Blood carries the lipoproteins to adipose cells for storage. The liver also produces phospholipids and cholesterol, which are needed for the hepatic production of bile salts, steroid hormones, components of cell membranes and other special molecules. Fats can also be produced (synthesised) from carbohydrate and protein, primarily in the liver. The role of lipids in the body is summarised in Table 26-2.
Table 26-2 THE IMPORTANCE OF PROTEINS, CARBOHYDRATES AND FATS IN THE BODY
| Proteins | |
| • | Contraction of muscle (actin and myosin are proteins) |
| • | Energy (proteins can be metabolised for energy) |
| • | Fluid balance (the protein albumin is a major source of plasma oncotic pressure) |
| • | Protection (antibodies and complement are proteins associated with immunity) |
| • | Regulation (enzymes regulate chemical reactions, while hormones regulate many physiological processes) |
| • | Structure (collagen fibres provide structural support to many parts of the body; keratin strengthens skin) |
| • | Transport (haemoglobin and plasma proteins are transport molecules, while proteins in cell membranes control movement of materials into and out of cells) |
| Carbohydrates | |
| • | Energy (glucose is the main energy source for most body cells; neurons and red blood cells are dependent on glucose) |
| • | Storage (energy stores in the liver are released into the bloodstream when needed) |
| Fats | |
| • | Structure (cell membranes are predominantly made of lipid molecules) |
| • | Insulation (myelination of neurons consists of lipids, while fat stores under the skin provide insulation from heat loss) |
| • | Protection (fatty deposits around soft tissues provide physical protection) |
| • | Storage (energy is stored in lipid molecules) |
Proteins
Protein synthesis (production) requires the presence of all the essential amino acids — obtained only from food — as well as nonessential amino acids (those that can be produced by the body). Proteins perform many important roles in the body; these are summarised in Table 26-2. Amino acids can be used for energy but first ammonia needs to be removed within the liver (known as deamination). Ammonia is toxic to the body and needs to be removed. It is converted to urea by the liver and passes into the blood to be excreted by the kidneys. Depending on need, ketoacids may be converted to fatty acids for fat synthesis and storage or used to provide energy for the liver cells.
The plasma proteins, including albumins and globulins (which are important proteins involved in transport, enzyme and fluid regulation) are produced by the liver. They play an important role in maintaining blood volume and pressure by maintaining plasma oncotic pressure (see Chapter 22 for more details). The liver also produces several nonessential amino acids and serum enzymes, including aspartate aminotransferase (AST), alanine aminotransferase (ALT), lactate dehydrogenase (LDH) and alkaline phosphatase (ALP). Common tests of liver function are listed in Table 26-3. When there is suspected damage to the liver or metabolic processes are altered (discussed in Chapter 27), liver function tests are often performed. These tests measure a broad range of hepatocyte and biliary functions to provide an overview of liver function.
Table 26-3 SELECTED TESTS OF LIVER FUNCTION
| TEST | NORMAL VALUE | CLINICAL SIGNIFICANCE |
|---|---|---|
| Serum enzymes | ||
| Alkaline phosphatase (ALP) | 25–100 U/L | Increases with biliary obstruction and cholestatic hepatitis |
| Aspartate aminotransferase (AST) | 0–40 U/L | Increases with hepatocellular injury |
| Alanine aminotransferase (ALT) | 0–35 U/L | Increases with hepatocellular injury |
| Lactic dehydrogenase (LDH) | 110–230 U/L | Elevated with hypoxic and primary liver injury |
| Gamma glutamyl transferase (GGT) | Females 0–30 U/L Males 0–50 U/L | Elevated with alcohol consumption (and cholestatic disease) |
| Bilirubin metabolism | ||
| Serum bilirubin total | 0–20 µmol/L | Increases with biliary obstruction, hepatocellular injury or obstruction, haemolysis (lysis of red blood cells) |
| Serum proteins | ||
| Albumin | 32–45 g/L | Reduced with hepatocellular injury |
| Globulin | 25–35 g/L | Increases with hepatitis |
| Blood-clotting functions | ||
| Prothrombin time (PT) | 10–14 sec | Increases with chronic liver disease (cirrhosis) or vitamin K deficiency |
| Activated partial thromboplastin time (aPTT) | 25–40 sec | Increases with severe liver disease or heparin therapy |
Source: RCPA manual. Royal College of Pathologists of Australasia, Version 5. 2009. Available at www.rcpamanual.edu.au/sections/pathologytestindex.asp?s=33, accessed July 2010; SydPath. The Pathology Service of St Vincent’s Hospital Sydney, Australia. Available at www.sydpath.stvincents.com.au/tests/InfSheets.htm, accessed July 2009.
Carbohydrates
The liver contributes to the stability of blood glucose levels by releasing glucose during hypoglycaemia (low blood glucose) and taking up glucose during hyperglycaemia (high blood glucose) and storing it as glycogen (glyconeogenesis) or converting it to fat. When glucose needs to be released into the blood to increase blood glucose levels, the two main options are:
The roles of carbohydrates are summarised in Table 26-2.
Storage of minerals and vitamins
The liver stores certain vitamins and minerals, including iron and copper, in times of excessive intake and releases them in times of need. The liver can store vitamins B12 and D for several months and vitamin A for several years. The liver also stores vitamins E and K. Iron is stored in the liver as ferritin, an iron–protein complex, and is released as required for red blood cell production.
Metabolism of bilirubin
Bilirubin is a by-product of the destruction of aged red blood cells. It gives bile a greenish-black colour, and in large amounts produces the yellow tinge of the skin and eyes due to jaundice (a yellowish colour due to deposition of bilirubin).
Aged red blood cells are taken up and destroyed by macrophages of the mononuclear phagocyte system, primarily in the spleen and liver. Within these cells, haemoglobin is separated into its component parts: haem and globin. The globin component is further degraded into its constituent amino acids, which are recycled to form new protein. The iron from the haem component can be recycled — by being either stored in the liver until required or used by the bone marrow to make new red blood cells. Another product derived from haem is bilirubin. The liver metabolises this bilirubin and secretes the waste products, which form the pigments of the bile. It is important to distinguish between bile salts, which assist fat digestion, and bile pigments, which contain the wastes of bilirubin. After being secreted by the bile into the duodenum, the bilirubin reaches the terminal ileum and colon, where it is further processed by bacteria and converted to urobilinogen. Most of the urobilinogen is absorbed into the bloodstream and then excreted in the urine, and a small amount is eliminated in faeces.
Metabolic detoxification
The liver metabolises substances produced by the body (such as hormones), as well as those that have been taken into the body (such as drugs), to make them less toxic or less biologically active. This process, called metabolic detoxification (or biotransformation), facilitates their intestinal and renal excretion. In this way alcohol, barbiturates, amphetamines, steroids and hormones (including oestrogens, aldosterone, antidiuretic hormone and testosterone) are metabolised or detoxified, preventing excessive accumulation and adverse effects.
Although metabolic detoxification is usually protective, sometimes the end products become toxins. For example, the end products of alcohol metabolism are acetaldehyde and hydrogen. Excessive intake of alcohol over a prolonged period causes these end products to damage hepatocytes. Acetaldehyde damages cellular mitochondria and the excess hydrogen promotes fat accumulation. Therefore, alcohol impairs the liver’s ability to function (discussed further in Chapter 27).
The gallbladder
The gallbladder is a saclike organ on the inferior surface of the liver. Its primary function is to store and concentrate bile between meals. Bile flows from the liver through the right or left hepatic duct into the common hepatic duct to the closed sphincter of Oddi; this controls flow into the duodenum and prevents backflow of duodenal contents into the pancreatobiliary system. Bile then flows through the cystic duct into the gallbladder, where it is concentrated and stored. The mucosa of the gallbladder wall readily absorbs water and electrolytes, leaving a high concentration of bile salts, bile pigments and cholesterol. The gallbladder holds about 90 mL of bile.
Within 30 minutes after eating, the gallbladder begins to contract and the sphincter of Oddi relaxes, forcing bile into the duodenum through the major duodenal papilla. During the cephalic and gastric phases of digestion, gallbladder contraction is facilitated by the parasympathetic nervous system (via branches of the vagus nerve). Hormonal regulation of gallbladder contraction is derived from the release of cholecystokinin and motilin secreted by the duodenal mucosa in the presence of fat. Vasoactive intestinal peptide, pancreatic polypeptide and sympathetic nerve stimulation relax the gallbladder.
The pancreas
The pancreas is approximately 20 cm long, with its head tucked into the curve of the duodenum and its tail touching the spleen. The body of the pancreas lies deep in the abdomen, behind the stomach (see Figure 26-24). The pancreas is unique in that it has both endocrine and exocrine functions. The endocrine function is to secrete hormones directly into the bloodstream: mainly insulin and glucagon. Pancreatic hormones are discussed in Chapter 10.
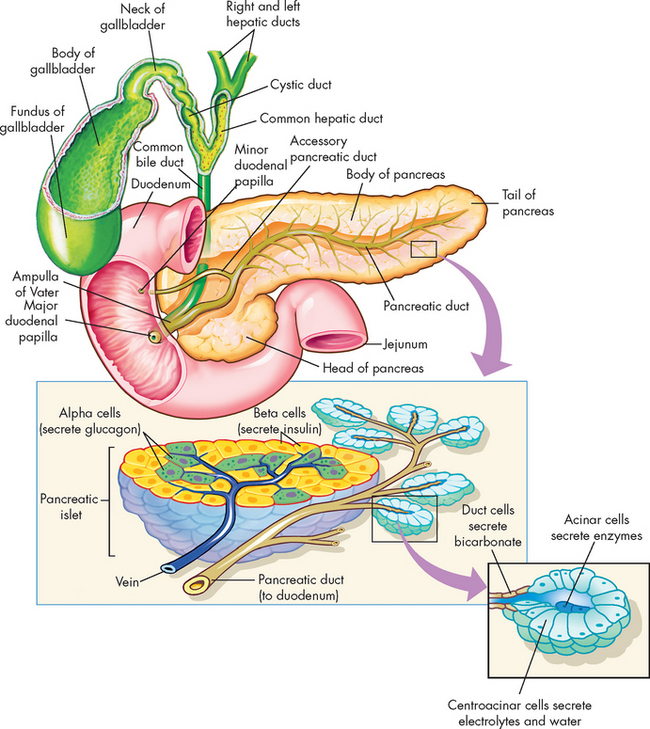
FIGURE 26-24 Associated structures of the gallbladder, pancreas and pancreatic acinar cells and duct.
Source: Main illustration from Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
The exocrine function consists of the secretions that empty directly into the digestive tract (and thus are not secreted into the bloodstream) — this is the function of the pancreas that we consider in this section on the gastrointestinal tract. The pancreas is composed of acini and networks of ducts that secrete enzymes and alkaline fluids with important digestive functions. The acinar cells are organised into spherical lobules around small secretory ducts (see Figure 26-24). Secretions drain into a system of ducts that leads to the pancreatic duct, which empties into the common bile duct at the ampulla of Vater.
The aqueous secretions of the exocrine pancreas contain potassium, sodium, bicarbonate and chloride. The highly alkaline (bicarbonate-rich) pancreatic juice neutralises the acidic chyme that enters the duodenum from the stomach and provides the alkaline medium needed for the actions of digestive enzymes and intestinal absorption of fat. Because eating stimulates the flow of pancreatic juice, the juice is most alkaline when it needs to be: during digestion.
The pancreas is a major source of enzymes for the gastrointestinal tract, as enzymes involved in the chemical digestion of the three main nutrients (proteins, fats and carbohydrates) arise from the pancreas. There are a few enzymes that digest protein, such as trypsin — this is produced in an inactive form, to protect the pancreas from the digestive effects of its own enzymes. Pancreatic lipase digests lipids (fats), while pancreatic amylase digests carbohydrates (remember, salivary amylase is also involved in carbohydrate digestion).
Secretion of the aqueous and enzymatic components of pancreatic juice is controlled by hormonal and vagal stimuli. Secretin stimulates the acinar and duct cells to secrete the bicarbonate-rich fluid that neutralises chyme and prepares it for enzymatic digestion. As chyme enters the duodenum, its acidity (pH of 4.5 or less) stimulates the S cells (secretin-producing cells) of the duodenum to release secretin, which is absorbed by the intestine and delivered to the pancreas in the bloodstream. In the pancreas, secretin causes the ductal and acinar cells to release alkaline fluid. Secretin also inhibits the actions of gastrin, thereby decreasing gastric acid secretion and motility. The overall effect is to neutralise the contents of the duodenum.
Enzymatic secretion follows, stimulated by cholecystokinin and acetylcholine. Cholecystokinin is released in the duodenum in response to the essential amino acids and fatty acids already present in chyme. Cholecystokinin and acetylcholine both act on the acinar cells, causing enzyme release. Once in the small intestine, activated pancreatic enzymes inhibit the release of more cholecystokinin and acetylcholine. This feedback mechanism inhibits the secretion of more pancreatic enzymes. Acetylcholine is liberated from pancreatic branches of the vagus nerve during the cephalic phase of digestion. Pancreatic polypeptide is released after eating and inhibits postprandial pancreatic exocrine secretion. (See Table 26-1 for a summary of hormonal stimulation of pancreatic secretions.)
THE LARGE INTESTINE
Anatomy and physiology of the large intestine
The large intestine (or large bowel) has a relatively large diameter of 7 cm, is approximately 1.5 m long and consists of the caecum, appendix, colon (ascending, transverse, descending and sigmoid), rectum and anal canal (see Figure 26-25). The caecum is a pouch that receives chyme from the ileum. Attached to it is the vermiform appendix, an appendage having only little function in immunity. From the caecum, chyme enters the colon, which loops upwards, traverses the abdominal cavity and descends to the anal canal. The four parts of the colon are the ascending colon, transverse colon, descending colon and sigmoid colon. Two sphincters control the flow of intestinal contents through the caecum and colon: the ileocaecal valve, which admits chyme from the ileum to the caecum, and the O’Beirne sphincter, which controls the movement of wastes from the sigmoid colon into the rectum. A thick (2.5–3 cm) portion of smooth muscle surrounds the anal canal, forming the internal anal sphincter. Overlapping it distally is the striated muscle of the external anal sphincter.
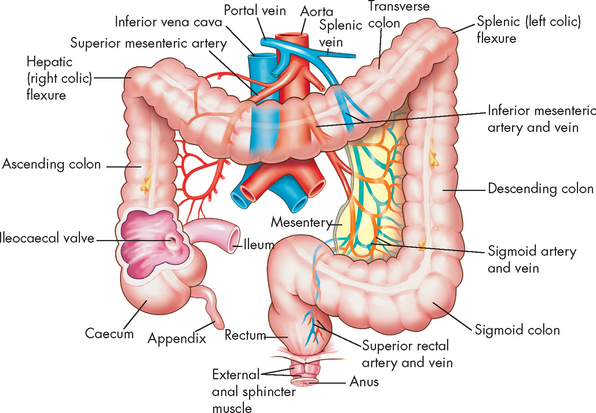
FIGURE 26-25 Division of the large intestine.
Source: Patton KT, Thibodeau GA. Anatomy & physiology. 7th edn. St Louis: Mosby; 2010.
In the caecum and colon, the longitudinal muscle layer consists of three longitudinal bands called teniae coli (see Figure 26-25). They are shorter than the colon and give it a gathered appearance. The circular muscles of the colon separate the gathers into outpouchings called haustra (plural; singular = haustrum). The haustra become more or less prominent with the contractions and relaxations of the circular muscles. The mucosal surface of the colon has rugae (folds), particularly between the haustra and Lieberkühn crypts, but no villi. Columnar epithelial cells and mucus-secreting goblet cells form the mucosa throughout the large intestine. The columnar epithelium absorbs fluid and electrolytes and the mucus-secreting cells lubricate the mucosa.
Most of the movements within the large intestine involve segmentation. The circular muscles contract and relax at different sites, shuffling the intestinal contents back and forth between the haustra, most commonly during fasting. The movements massage the intestinal contents, called the faecal mass at that point, and facilitate the absorption of water. Propulsive movement occurs with the proximal-to-distal contraction of several haustral units. Peristaltic movements also occur and promote the emptying of the colon — these mass movements occur a few times per day. The gastrocolic reflex initiates propulsion in the entire colon, usually during or immediately after eating, when chyme enters from the ileum. This reflex causes the faecal mass to pass rapidly into the sigmoid colon and rectum, stimulating defecation. Gastrin may participate in stimulating this reflex.
Intestinal bacteria
The number of bacteria increases from the stomach to the distal colon. The stomach is relatively sterile because of the secretion of acid that kills ingested pathogens or inhibits bacterial growth. Bile acid secretion, intestinal motility and antibody production suppress bacterial growth in the duodenum, and in the duodenum and jejunum there is a low concentration of aerobes, primarily streptococci, lactobacilli, staphylococci and enterobacteria. Anaerobes are found distal to the ileocaecal valve but not proximal to the ileum. They constitute about 95% of the faecal flora in the colon and contribute one-third of the solid bulk of faeces. Bacteroides, clostridia, anaerobic lactobacilli and coliforms are the most common microorganisms from the ileum to the caecum.
The intestinal bacteria survive on the indigestible carbohydrates (fibre) within the large intestine. They ferment these carbohydrates, releasing large amounts of gases (some of which exits the body via the rectum, known as flatulence). The bacteria perform some functions that are beneficial to the human host, including releasing some vitamins (some of the B group vitamins, as well as vitamin K), which can then be absorbed into the bloodstream and contribution to nutrition. The bacteria also have a role in the metabolism of bile salts, oestrogens, androgens, lipids, various nitrogenous substances and drugs, and protection against infection. The normal flora do not have the virulence factors associated with pathogenic microorganisms, thus permitting immune tolerance.5 Infections of the lower gastrointestinal tract occur by three major mechanisms: proliferation or overgrowth of bacteria, perforation of the intestine and contamination of neighbouring structures.
Defecation reflex
The movement of faeces into the sigmoid colon and rectum stimulates the defecation reflex (rectal reflex). The rectal wall stretches and the constricted internal anal sphincter (smooth muscle with autonomic nervous system control) relaxes, creating the urge to defecate (see Figure 26-26). The defecation reflex can be overridden voluntarily by contraction of the external anal sphincter and muscles of the pelvic floor. The rectal wall gradually relaxes, reducing tension and the urge to defecate passes. Retrograde contraction of the rectum may displace the faeces out of the rectal vault until a more convenient time for evacuation. Pain or fear of pain associated with defecation (for example, due to haemorrhoids) can inhibit the defecation reflex.
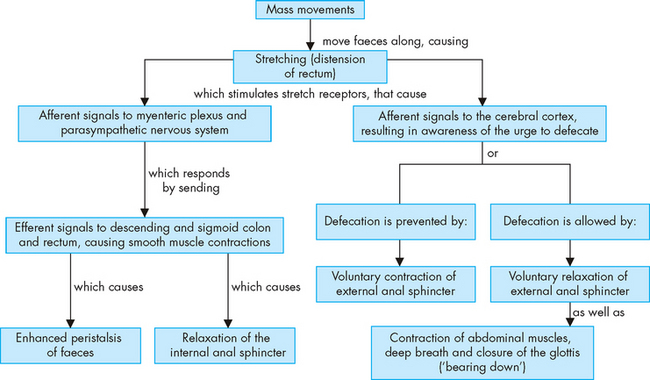
Squatting and sitting facilitate defecation because these positions straighten the angle between the rectum and anal canal and increase the efficiency of straining (increasing intra-abdominal pressure). Intra-abdominal pressure is increased by initiating the Valsalva manoeuvre — that is, inhaling and forcing the diaphragm and chest muscles against the closed glottis to increase both intrathoracic and intra-abdominal pressure, which is transmitted to the rectum.
Rectal valves are projections of tissue into the lumen of the rectum — these valves have the important function of helping to hold the faeces, while allowing gases to escape.
Digestion in the large intestine
The majority of nutrient absorption has already occurred in the small intestine, so there is no substantial digestion remaining in the large intestine. Although the bacteria of the large intestine digest the carbohydrates as their own fuel source, this does not provide carbohydrates for the human host.
Absorption in the large intestine
The large intestine absorbs water and some fatty acids (produced by bacterial fermentation). Absorption occurs in the caecum, ascending colon, transverse colon and descending colon. Vitamins B and K (produced by the bacteria) are also absorbed here. By the time the faecal mass enters the sigmoid colon, the mass consists entirely of wastes and is called faeces (or stools), consisting of food residue, unabsorbed gastrointestinal secretions, shed epithelial cells and bacteria. Absorption of some drugs can occur by rectal administration, whereby the drugs are absorbed quickly into the bloodstream.
Fluid movements in the digestive system
Approximately 8.5 L of fluid enters the digestive system every day (see Figure 26-27): 2 L is ingested and the remaining 6.5 L consists of secretions (from the saliva, stomach, bile, pancreas and small intestine). Of this volume, 99% is absorbed: 90% (7.6 L) in the small intestine and 9% (0.8 L) in the colon. The remaining 100 mL of water is excreted daily in the faeces. The amount of fluid within the digestive system, especially the lower intestine, is important. Diarrhoea causes large water loss and can lead to dehydration, particularly in the young and the elderly.
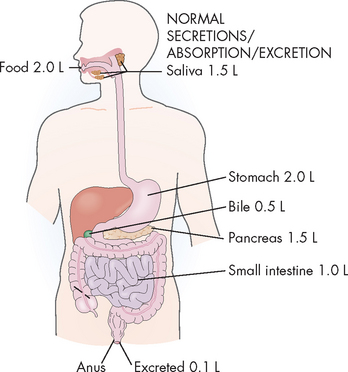
FIGURE 26-27 Normal intake, absorption, secretion and excretion of water.
Source: Damjanov I. Pathophysiology. Philadelphia: Saunders; 2009.
AN OVERVIEW OF NUTRITION
The main nutrients that the body needs are proteins, carbohydrates and fats. In addition, water needs to be consumed in relatively large amounts. Water is discussed fully in Chapters 28 and 29. In this section, we briefly examine human nutrition.
Protein is found in highest amounts in animal products; sources of protein in the diet include meat, fish, poultry, eggs, dairy and cereals. Protein is digested and broken down into amino acids, which are then processed by the liver to produce new proteins that have a wide variety of uses in the human body (see Table 26-2).
Dietary carbohydrates are mainly of plant origin. Starch (a complex carbohydrate or polysaccharide) is found in cereals, grains, fruits and vegetables. Fibre (or non-starch polysaccharide) is a type of carbohydrate that we are unable to break down and absorb, and it has an important role in maintaining the health of the large intestine (see Chapter 27). Sources of dietary fibre include wholemeal and wholegrain breads and cereals, oats, fruits and vegetables. The sugars, or simple carbohydrates, are not often found in natural foods, other than fructose in honey; they are used in the production of processed foods, such as soft drinks, ice-cream, cakes and biscuits. Carbohydrates are digested and broken down into simple sugars (monosaccharides), namely glucose, galactose and fructose. These are used as an energy source in the form of glucose (see Table 26-2).
Fats (or lipids) in the diet are mainly in the form of triglycerides, although we also consume other types such as cholesterol and phospholipids. Different chemical structures of fats include saturated fats (from animal sources, such as butter, cream and fat in meat), monounsaturated fats (usually from plant sources, such as olive oil) and polyunsaturated fats (further subdivided into types including omega-3 [from fish] and omega-6 [from sesame, linseed and peanut oils]). Trans fatty acids are most prominent in our diet in margarines. Further details of some types of dietary fats are given in Box 26-2. Fats are broken down to triglycerides, and small lipid molecules are processed by the liver for a number of functions in the body (see Table 26-2).
Energy intake that is beyond the needs of the body is stored for potential usage later. In Australia and New Zealand, more than half the population are overweight and obese — a serious health concern in modern times, due to imbalance in energy intake and output. A detailed discussion of this issue can be found in Chapter 35.
Although vitamins (organic) and minerals (inorganic) are necessary in only very small amounts, they are nevertheless important for adequate health. They are absorbed into the blood without being digested to smaller components. A full discussion of all vitamins and minerals is beyond the scope of this text, but Table 26-4 lists those for which we have dietary guidelines in Australia and New Zealand.
Table 26-4 NUTRIENTS FOR WHICH WE HAVE DIETARY GUIDELINES IN AUSTRALIA AND NEW ZEALAND
| MAIN NUTRIENTS | VITAMINS | MINERALS |
|---|---|---|
Source: Australian Government Department of Health and Ageing, National Health and Medical Research Council, Ministry of Health. Nutrient reference values for Australia and New Zealand (2006).
In recent years, studies of chronic disease have indicated that modifying our intake of particular nutrients can help to promote health. For example, eating sufficient amounts of fruits and vegetables can prevent ischaemic heart disease and a number of different cancers.6 Accordingly, dietary guidelines are also available for modifying food intake to decrease the risk of developing chronic diseases (see Table 26-5).
Table 26-5 ADAPTING NUTRIENT INTAKE TO REDUCE THE RISK OF CHRONIC DISEASE
| TO REDUCE THE RISK OF CHRONIC DISEASE | |
|---|---|
| INCREASE INTAKE OF: | DECREASE INTAKE OF: |
Source: Australian Government Department of Health and Ageing, National Health and Medical Research Council, Ministry of Health. Nutrient reference values for Australia and New Zealand (2006).
AGEING AND THE DIGESTIVE SYSTEM
With increasing age, the gastrointestinal system experiences a progressive decline in normal function.7–10 8 9 10 These changes can lead to a decline in the quantity and variety of food that is ingested, digested and absorbed, leading to poorer nutrition for the individual.
Teeth are lost as a result of periodontal disease and brittle roots that break easily. Taste buds decline in number and the sense of smell diminishes. Salivary secretion decreases, resulting in a dry mouth. Dysphagia (difficulty swallowing, discussed fully in Chapter 27) becomes more common, particularly in those who have suffered from stroke. As a result of these changes, eating is less pleasurable, appetite is reduced and food is not chewed or lubricated enough, so swallowing is difficult.
Gastric motility, blood flow, and the volume and acid content of gastric juice may be reduced, particularly with gastric atrophy (shrinkage of stomach tissue). The decreased gastric secretions can impair absorption of vitamin B12, calcium, iron and folate. The protective mucosal barrier decreases, so there is increased susceptibility to damage. There are decreases in intestinal absorption, motility and blood flow, which result in nutritive substances being absorbed more slowly and in smaller amounts. The rectal muscle mass decreases and the anal sphincter weakens. Constipation is common and is related to lowered intestinal motility, immobility, a low-fibre diet and changes in enteric nervous system functions.
The overall size and weight of the liver decreases. Both the blood flow and the enzyme activity of the liver decrease, so the ability to detoxify substances such as drugs decreases. In the pancreas, fibrosis, fatty acid deposits and atrophy occurs. There is a corresponding decrease in pancreatic secretion of digestive enzymes, particularly those that digest proteins. Although there are no obvious changes in the gallbladder and bile ducts, there is an increased prevalence of gallstones and cholecystitis (see Chapter 27).
An overview of the digestive system
The gastrointestinal tract is a hollow tube that extends from the mouth to the anus and is divided into several organs: the mouth, pharynx, oesophagus, stomach, small intestine and large intestine. The salivary glands, liver, gallbladder and pancreas are the main accessory organs of this system. The major functions of the gastrointestinal tract are the mechanical and chemical breakdown of food and the absorption of digested nutrients into the bloodstream. The walls of the gastrointestinal tract have four layers: mucosa, submucosa, muscularis (circular muscle and longitudinal muscle) and serosa. The main nervous system control of the digestive system is by the enteric nervous system, which is located within the gut wall. In addition, the parasympathetic nervous system (via the vagus nerve) stimulates gastrointestinal motility and sphincter relaxation; and the sympathetic nervous system decreases gastrointestinal activity.The mouth, pharynx and oesophagus
Digestion begins in the mouth with chewing and salivation. Salivary amylase initiates carbohydrate digestion. The oesophagus is a muscular tube that transports food from the mouth to the stomach. The tunica muscularis in the upper part of the oesophagus is striated muscle and that in the lower part is smooth muscle.The stomach
The stomach secretes digestive juices, mixes and stores food, and propels partially digested food (chyme) into the duodenum. The hormones gastrin and motilin stimulate gastric emptying; the hormones secretin and cholecystokinin delay gastric emptying. Mucus is secreted throughout the stomach and protects the stomach wall from acid and digestive enzymes. Gastric glands in the fundus and body of the stomach secrete intrinsic factor (needed for vitamin B12 absorption) and hydrochloric acid, which dissolves food fibres, kills microorganisms and activates the enzyme pepsin. Chief cells in the stomach secrete pepsinogen, which is converted to pepsin in the acid environment created by hydrochloric acid. Acid secretion is stimulated by the vagus nerve and gastrin, and is inhibited by sympathetic stimulation and cholecystokinin.The small intestine
The peritoneum is a double layer of membranous tissue: the visceral layer covers the abdominal organs and the parietal layer extends along the abdominal wall. Villi are small finger-like projections that extend from the small intestinal mucosa and increase its absorptive surface area. Digested substances are absorbed across the intestinal wall and then transported to the liver, where they are metabolised further. The majority of all absorption occurs in the small intestine. Small molecules of carbohydrates, proteins and lipids are absorbed primarily by the duodenum and jejunum; bile salts and vitamin B12 are absorbed by the ileum. Vitamin B12 absorption requires the presence of intrinsic factor.Accessory organs of digestion
There are two blood vessels into the liver: the hepatic portal vein and the hepatic artery. All blood exiting the liver travels through the hepatic vein The hepatocytes produce 700–1200 mL of bile per day and secrete it into the bile canaliculi, which are small channels between the hepatocytes. The bile canaliculi drain bile into the common bile duct and then into the duodenum through an opening called the sphincter of Oddi. Sinusoids are capillaries located between the plates of hepatocytes. Blood from the portal vein and hepatic artery flows through the sinusoids to a central vein in each lobule and then to the hepatic vein and inferior vena cava. Kupffer cells, which are part of the mononuclear phagocyte system, line the sinusoids and destroy microorganisms in sinusoidal blood. Bile is produced by the liver and is necessary for fat digestion and absorption. Bile’s alkalinity helps to neutralise chyme, thereby creating a pH that enables the pancreatic enzymes to digest proteins, carbohydrates and sugars. Bile salts enable fats to be emulsified to reach the brush border of the intestinal mucosa. The fat content of the micelles diffuses through the epithelium into lacteals (lymphatic ducts) in the villi. From there, fats flow into the lymphatics and into the systemic circulation. Most bile salts and acids are recycled. The absorption of bile salts and acids from the terminal ileum and their return to the liver is known as the enterohepatic circulation of bile. The liver has an important role in the metabolism of the main nutrients, such that they can be used in the production of new molecules and excesses may be stored until needed. Bilirubin is a pigment released from the lysis of aged red blood cells. The liver metabolises the bilirubin and it is then secreted with the bile as bile pigments. This metabolised bilirubin is excreted from the body in the faeces. The gallbladder is a saclike organ located in the inferior surface of the liver. The gallbladder stores bile between meals and ejects it when chyme enters the duodenum. Stimulated by cholecystokinin, the gallbladder contracts and forces bile through the cystic duct and into the common bile duct. The sphincter of Oddi relaxes, enabling bile to flow through the major duodenal papilla into the duodenum. The pancreas is an organ located behind the stomach. The pancreatic secretions for digestion include an alkaline solution and enzymes that digest proteins, carbohydrates and fats. Secretin stimulates pancreatic secretion of alkaline fluid, and cholecystokinin and acetylcholine stimulate secretion of enzymes. Pancreatic secretions originate in acini and ducts of the pancreas and empty into the duodenum through the common bile duct or an accessory duct that opens directly into the duodenum.The large intestine
The large intestine consists of the caecum, appendix, colon (ascending, transverse, descending and sigmoid), rectum and anal canal. Haustra are pouches of colon formed with alternating contraction and relaxation of the circular muscles. Distention of the ileum with chyme causes the gastrocolic reflex, or the mass propulsion of faeces to the rectum. Defecation is stimulated when the rectum is distended with faeces. The contracted internal anal sphincter relaxes and if the voluntarily regulated external sphincter relaxes, defecation occurs. The largest number of intestinal bacteria are in the colon. They are anaerobes consisting of Bacteroides, clostridia, coliforms and lactobacilli. Intestinal bacteria digest carbohydrates and produce vitamins that can then be absorbed through the intestinal wall and into the bloodstream.An overview of nutrition
Rich sources of protein are found in animal products. Proteins are digested to amino acids and then used by the body for a wide variety of structural and functional purposes. Carbohydrates are mainly found in plants. Starch is digested to simple sugars, which are mainly used by the body as an energy source in the form of glucose. Fibre is a non-digestible carbohydrate, which assists with progressing intestinal contents through at an appropriate rate.Emily is a 34-year-old librarian. She has just had her lunch break, during which she ate a chicken and salad burger with hot chips at a nearby café and sat quietly reading a book while eating. After that, she ate a large slice of chocolate cake, which she had difficulty finishing. Although Emily enjoyed her lunch, on returning to work she feels tired and wishes she could have a nap.